a. Primates
Living organisms represent a small part of all species that have lived in the past. Many species have disappeared due to catastrophic events or competition with other species, and their fossils are preserved in the geologic record. George Cuvier (1769-1832) [Essay on the Theory of the Earth, 1813] noticed that many organisms had not survived to recent times and are only represented in the fossil record. Cuvier identified five major massive extinction events that decimated the number of living species (Fig. 1), of which the fifth (the K/T, Cretaceous-Tertiary, transition) was caused by the impact of an asteroid in the Yucatan peninsula (65 Ma) causing the Chicxulub crater, a 180 km-diameter ring (https://en.wikipedia.org/wiki/Chicxulub_crater). This massive extinction represented a decrease of over 50% of living organisms during the Permian (3). The Dinosaurs went extinct allowing mammals to diversify.
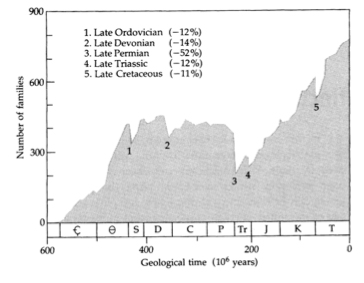
Figure 1. Major extinctions in the fossil record.
The evidence of massive extinctions of organisms underlies the geologic time-scale classification. Primates greatly diversified during the Tertiary period, and the hominines diverged during the Pliocene. This time-line classification is useful for defining the evolutionary landmarks of our ancestors. Mammal differentiation dates back to the Triassic/Jurassic transition, 206 Ma (Fig. 2). Triassic mammal-like reptiles (Therapsids) share with mammals a unique trait: heterodont dentitions. Modern amphibians and reptiles have homodondt teeth; all their teeth show a conical shape wit a single, projecting point or cusp. This primitive condition serves to capture preys but not for food processing or chewing. Therapsids show some anatomically differentiated teeth, such as a projecting canine and a non-single cusped tooth that can be called an incipient molar tooth. Therapsids heterodontia was likely due to a shift in dietary habits that required a diversification of the functional use of teeth.
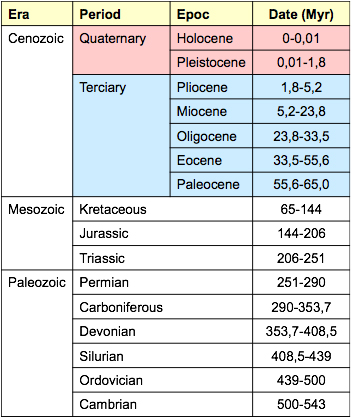
Figure 2. Geologic periods based on major extinction events.
Castrocauda is one of the most ancient mammalian fossils [Science 311, 1123-1127, 2006]. Dated to 164 Ma, in the Jurassic period, it shows evidence of a hair fur covering the body (hair predates many other derived traits of modern mammals) and a tail adapted to aquatic environments, similar to modern Ornithorhynchus. The docodonts include late Mesozoic mammaliform organisms as Castrocauda, whose most distinguishing dental feature was their relatively sophisticated molar teeth. Castrocauda shows that these animals were not all tiny insectivorous shrew-like organisms, but also highly adapted, despite the reptilian domination.
Repenomamus robustus and R. giganticus were terrestrial mammaliforms from deposits in China that date to the early Cretaceous period, 125-123.2 Ma. There is good evidence that R. robustus fed on small vertebrates and young dinosaurs, either by scavenging or opportunistic hunting, while R. giganticus [Nature, 433, 149-152, 2005], a large mammal from the Mesozoic era, had a dental formula (3124/2125) with mixed primitive (5 molars) and derived (2 premolars) traits.
Primate diversification within the mammals, goes back at least 65 Ma. Molecular analyses have estimated the origin of the primate branch in the mid-Cretaceous period, around 85 Ma, well before the extinction of the dinosaurs. However, one of the oldest known primates from the fossil record is Altiatlasius, dated to the Late Paleocene in Africa and to the Paleocene–Eocene transition in the northern continents. Other remains of ancient primates date to 55 Ma, such as Cantius, Donrussellia, and Altanius. More recent specimens are Teilhardina, a well-known, early Eocene (56-47 Ma) marmoset-like primate from Europe, North America and Asia, and Darwinius masillae, a species within the adapoidea, a 47 Ma basal strepsirhini from the middle Eocene, representative of the early haplorrhini diversification. Another significant fossil is Eosimias, a 40-45 Ma middle Eocene specimen between simians and prosimians, close to the phylogenetic position of the tarsiers.
Within the order of primates (O. Primates) we shall include placental mammals that always have a clavicle bone in the shoulder girdle, nails in at least one finger instead of claws (if not in all fingers), a complete osseous ring surrounding the orbit, which may not be fully separated from temporal fossa, a trend to show the orbits in a ventral (frontal) position, instead of lateralised, and a variable degree of opposable thumbs. Primates always have a well-developed Calcarine fissure that defines the posterior lobe of the brain. Many of these anatomical traits greatly vary within the primates, especially between the prosimians and anthropoids.

The Sb.O. Strepsirrhini includes primates in which the orbital and temporal fossae are clearly communicated and with an orbital rim defining the border of the orbits. In this trait they differ from the insectivorous mammals that do not have a postorbital bar and from the haplorhines that have a fully closed orbit (Fig. 3). The prosimians have unfused hemi-mandibular bones, a humid rhinarium lacking hair, and laterally oriented nostrils (nasal holes). Three Sp.F. are recognized within the prosimians: the Lemuroidea, Daubentonioidea and Lorisoidea. Most prosimians are nocturnal animals with large eyes, somewhat laterally placed, and quick sexual maturation; they frequently give birth to several offspring, have two or more pairs of mammary glands and a bicornic uterus; their face is long, with a prominent and projected face; their upper lip is fused to the gum, the dental formula varies from 3143 to 3123 and 2123, so they may have 2 or 3 incisors, and from 4 (the most primitive condition) to 2 premolars (the most derived one), the incisors and canine are generally projected to form a toothcomb, and most retain a claw in each foot for cleaning purposes. Most prosimians live in Madagascar and Comores Islands, though the galagos also inhabit east Africa. They occupy arboreal habitats and feed on leaves, fruits and mainly on insects. Their hind limbs are longer that the forearms evidencing their mainly jumping locomotion. The Daubentonioidea include several highly specialised species that share a derived dental condition and dietary specializations.
The Sb.O. Haplorhini includes all the anthropoid primates (platyrrhines and catarrhines) and the Infr.O. Tarsiform, formerly included in the prosimians. They show fully closed orbits and hemimandibles (except in Tarsius that has partially closed orbits and non fused mandibular bones), rounded nostrils, and a non-humid nose, mostly with hair. The Tarsioidea do not have a humid rhinarium, which differs from the prosimians, but their postorbital closure is partially open (Fig. 3). They show large eyes, adapted to a nocturnal lifestyle, an insectivorous diet, absence of toothcomb, two claws at each foot for hair combing and long tarsal bone. The other anthropoidea primates show lager skulls, mobile upper lip, non-humid nose with hair, frontally placed orbits, fully separated from the temporal fossa, and anterior limbs longer than hind limbs. The Infr.O. Platyrhini (New World monkeys) show a flat and broad nose and nasal holes separated by a cartilaginous septum. They include two families: Cebidae (tree dwelling primates with prehensile tail) and Callithricidae (small, insectivorous primates with claws and absence of third molar). The Infr.O. Catarrhini (Old World monkeys) show a long snout, sciatic callosities, mostly terrestrial quadruped locomotion, and complex social behaviour. The old world monkeys include three superfamilies: Propliopithecidae, a fossil group including the genus Aegiptopithecus; Cercopithecoidea, including the macaques (Sb.F. Cercopithecinae) and langurs (Sb.F. Colobinae), and Hominoidea, that includes hylobatids, pongids and hominids. Humans are included into the F. Hominidae along with chimpanzees and gorillas. However, they are clearly separated from the orangutans (F. Pongidae) and the gibbons (F. Hylobatidae).
The cercopithecoidea and hominoidea are considered sister taxa. Compared to cercopithecoidea, the hominoidea include large bodied, diurnal primates that live in forested environments, have a greatly variable dietary regimens, from folivorous or frugivorous to omnivorous, display a mainly brachiator locomotion, though also terrestrial quadrupedal or even bipedal, and show a overall polyginic mode of reproduction. Some anatomical traits of the hominoidea can be considered primitive (plesiomorphic) while others are highly specialized (apomorphic). Their most significant derived anatomical traits are a large brain, reduced facial prognatism, wide thorax with a dorsally placed scapula, long forelimbs with a short olecranon, reduced lumbar region, wide sacrum, and absence of a tail. They also have significant life history and behavioural specializations, such as long gestation period, long infancy, greater reliance on learned behaviours, and greater dependence of infants. The hominoidea show few but significant non-specialized traits that resemble those seen in the anthropoid ancestors: patrilineal societies, a tubular tympanic bone, 2123 (2 incisors, 1 canine, 2 premolars, 3 molars) dental formula, and a bunodont dentition. The hominoidea include three distinct extant families: hylobatidae, pongidae and hominidae.
Hylobatidae. The hylobatids (gibbons) include several species of the Hylobates genus (H. syndactilus, H. concolor, H. hoolock, and H. lar). They live in Asia (India, China, Borneo, Java, and Sumatra). Their main characteristic traits are: 5-10 Kg, lack of sexual dimorphism, short face, absence of neurocranium crests, wide mandibular branch, short and wide incisors, long and prominent canines in both sexes, sectorial P3, bicuspid P4, low crowned molars with rounded cusps (bunodont), long arms (intermembral index of 126-147), and presence of reduced sciatic callosities. The hylobatids inhabit humid forests and display specialized suspensory brachiation locomotion; eat tender leaves, ripe fruits and insects. The occupy relatively small territories, where food items are dispersed, that include monogamous families of up to 4 offspring, giving birth a single new-born every 4-5 years. The male takes care of the infant during the second year of age and the infants remain within the family group until they are around 10 years old.
Pongidae. The pongids (orangutans) include a single species: Pongo pygmaeus. They live in Southeast Asia (Borneo, Sumatra), show a significant sexual dimorphism in size (females weigh 35 Kg on the average whereas mail weigh 78 Kg). They show vertical faces but with a significant alveolar prognatism and display large neurocranium crests, more pronounced in males; a wide mandibular branch, large central (I1) and small lateral (I2) upper incisors, sectorial P3 and bicuspid P4, low molars with rounded cusps and marked crests, and thick enamelled teeth; long arms (intermembral index of 139), short legs with reduced thumb, mainly suspensory brachiator but also terrestrial locomotion; they eat fruits, seeds, leafs, and bark. Males are solitary individuals while females live with the immature offspring until maturation occurs (around 7 years old in females and 8-15 in males. The male does not take care of infants and occupies and defends a large territory (against other males) where several females may inhabit.
Hominidae. This family includes gorillas, chimpanzees and humans, because molecular data have shown that the three taxa share a closer common ancestor than that shared with the orangutans and gibbons. All three inhabit African forested and woodland environments but show significant anatomical and behavioural differences.
Sub.F. Gorillinae. The number of species of gorillas has varied. At present two species are widely accepted, including several geographic subspecies: G. gorilla (G. gorilla gorilla or western gorilla, and G. gorilla diehli or north-western cross river gorilla) and G. beringei (eastern gorillas, including two subspecies: G. beringei beringei or mountain gorilla, and G. beringei graueri or eastern lowland gorilla). The gorillas show a great sexual dimorphism in size (females weight varies between 71 and 97 Kg, and males vary between 170 and 200 Kg), very wide and robust maxillary bones, marked cranial crests, and a protruding face. The incisors are small, whereas the canines, premolars and molars are large and very dimorphic. The have somewhat large arms (intermembral index of 116), short legs, and long thumbs. They live in green forests and mainly eat leafs, fruit when available, and bark. Males are mainly terrestrial, though young involve in bellow-branch locomotion. During terrestrial locomotion they practice knuckle-walking (placing the dorsal side of the second phalanx of the hands on the ground). Gorilla’s societies are polyginic with a single adult male and several non-related females moving throughout a large territory seeking dispersed foods. Males engage in competition for females forming harems, while young males may remain in the group and mature females show exogamous mating.
Sub.F. Homininae. This subfamily includes both chimpanzees and humans since they show significant molecular apomorphies indicative of a close common ancestor.
Tribe Panini. Since 1928 two distinct species have been recognized: the common chimpanzee (P. troglodytes) live north of the Congo River and the bonobo (P. paniscus) who live south. Pan troglodytes is divided into four subspecies (Pan troglodytes troglodytes, Pan troglodytes verus, Pan troglodytes ellioti, Pan troglodytes schweinfurthii [eastern chimpanzee], Pan troglodytes marungensis [southeastern chimpanzee]), while P. paniscus has none. Based on genome sequencing, the two extant Pan species diverged around one million years ago. The most obvious differences are that chimpanzees are somewhat larger, more aggressive and male dominated, while the bonobos are more gracile, peaceful, and female dominated. The common chimpanzees live in African tropical and arid savannah environments. They show reduced sexual dimorphism, with males weighing between 42 and 60 Kg, while females weigh between 33 and 45 Kg. They show a short face and reduced mandibular branch, wide incisors, moderate canine size, dimorphic between sexes, short premolars, and low molar crowns with rounded cusps (bunodont). They are both braquiators and terrestrial dwellers (knuckle-walking), with an intermembral index of 103-106 , showing somewhat short legs. They are mainly fruit eaters though also hunt small animals. Chimpanzee’s societies are fusion-fission structured depending on the availability of food resources. Kin related male groups defend a territory and a group of unrelated females that practice exogamy with other groups. The bonobo or pigmy chimpanzee weigh between 33 and 45 Kg and show little sexual dimorphism, restricted to the canine, Lives in somewhat closed environments and practice either braquiation, knuckle-walking, and limited bipedalism. Their social structure is not structured and sexual practices are used to prevent competition.
Tribe Hominini. The human lineage has greatly varied since the divergence from its LCA (last common ancestor) with the chimpanzee some 5.5 to 6 Ma. Modern humans show a significant sexual dimorphism. Males weigh 85 Kg on average and females 60 Kg. Human canines are greatly reduced compared to the chimpanzee and teeth show thick enamel, unlike gorillas and chimpanzees, but similar to the orangutan. Their face is flat, non-prognatic, and has a large brain, lacking crests on the skull. Human intermembral index is smaller that 100, showing long legs indicative of a bipedal locomotion. Their hallux is very long compared to the other apes. Human diet is omnivorous, including hunted meat. Social groups are highly structured and male competition for territory and reproduction are significant.
There are three main anatomical traits that need to be considered in the light of the reconstruction of the evolutionary process of the human lineage: 1) the origin of bipedal locomotion, 2) the functional significance of teeth and the anatomical structures associated to food chewing, and 3) the evolution of the brain. It was initially considered that intelligence was the first distinctive trait of humans, followed by bipedal locomotion and then the reduction of the masticatory apparatus. However, we now know that the first hominins attained bipedalism before having a large brain. Nevertheless, the first distinctive trait of the earliest hominins was the reduction of the dentition.
b. Bipedalism
The locomotor systems seen in primates are quadruped, cling and leap, brachiation, and bipedalism. Limb proportions (the ratio of the forelimb to hindlimb length) reflect the type of locomotion. Longer legs than arms are characteristic of jumpers, whereas quadrupeds tend to have somewhat shorter arms than legs and brachiators have longer arms than legs. The prosimians are mainly above-branch quadrupeds and jumpers, and most anthropoids are terrestrial quadrupeds, arboreal jumpers or bellow-branch brachiators. Humans differ from these patterns and are obliged bipeds. Some reptiles and the birds also walk on their hindlimb, although only the penguin has a vertically placed vertebral column while walking as humans do. Penguins are good at swimming, though they are clumsy on land. Human bipedalism is an efficient adaptation to travel long distances in open environments, since the human walk requires less energy than running and than quadrupedalism. During human bipedal march the body is in an unstable equilibrium in which the gravity center remains close to the vertebral column and the pelvic bone holds most of the body weight. During each step, the centre of gravity moves forward and a new step is required to prevent the body from falling down. In contrast, if a chimpanzee stands up, its center of gravity is above the pelvic bone and the vertebral column is placed behind the body axis, which requires a continuous use of muscles in the pelvic area to compensate the inefficient equilibrium.

There are many anatomical adaptations related to human bipedalism. In humans, the occipital hole (foramen magnum) in the occipital bone is more forward placed in the skull than in non-humans hominoids, as well as the articular condyles of the occipital bone that articulate with the first cervical vertebra, allowing the skull to remain in equilibrium without the need of a significant strength of the nucal muscles. The nucal insertion area of the skull and the spinal processes of the cervical vertebra are poorly developed in humans. In contrast, the gorillas show highly developed spinal processes in their cervical vertebrae to hold their strong nucal muscles. The size of the body of the vertebrae increases distally: it is larger in the lumbar vertebra to sustain the weight of the trunk. In addition, the human vertebral column is not straight; it shows two ventral curvatures (kyphosis), cervical and lumbar, and one dorsal curvature (lordosis). This allows a more stable position of the vertebral column without strong muscle and ligament insertions and a lowering of the center of gravity. The hominoids have a reduced mobility of the vertebral column compared to the cercopitecoids, including a reduction of the number of lumbar vertebrae, a common trait in brachiators that increases the speed of the swing movements while practicing bellow-branch brachiation. The human pelvis is wider and shorter than that seen in non-human hominoids, caused by a descent of the sacrum-iliac joint, which is also responsible for the descent of the gravity center. The bipedal locomotion also forces a rotation of the iliac crests from a dorsal to a lateral position, which allows the muscle gluteus minimum and gluteus medium (that go from the iliac crest to major trochanter on the femur) to perform an abductor function (a lateral separation of the leg), whereas in non-human hominoids they make an adductor function (extension) of the leg. Therefore, these two muscles help to maintain the lateral equilibrium during the bipedal gait. As a consequence of the rotation of the iliac crest, the sacrospinous ligaments (that go from the superior and inferior posterior spines to the sacrum, where they were associated to the movement of the tail) invade the pelvic canal thus reducing the size of the birth canal in humans. As a consequence of the reorganization of the vertebral column and the pelvic bones, the arms and legs also adapt to the bipedal locomotion. The gleno-humeral joint in the arm also moves laterally and the humerus experiments a torsion that allows the ventral to dorsal swing of the arm, reducing the ability to brachiate but allowing the manipulation of objects in the ventral region of the body. The femur in the leg also adapts to bipedalism by increasing the inward inclination of the femoral shaft to about 11º (4-5º in chimpanzees and 6º in gorillas), which compensates the separation of the femoral heads in a wide pelvis and reduces the separation of the knees while walking . The articular facets of the tibia are more excavated in humans, to reduce mobility of the knee, and the human foot is more robust than those of non-biped hominoids. The calcaneus bone is long and acts as a tripod with respect to the toes, and the big toe is in line with the rest of fingers, loosing its prehensile capabilities.
c. Maxillary bones and teeth
Teeth constitute the hardest structures of the human skeleton and, thus, they are differentially preserved in the fossil record. Teeth reflect the genetic basis since they are little affected by environmental conditions during growth and development and remain unchanged, except for dental wear, during life span. Teeth and bone structures associated to food chewing reflect long-term, evolutionary adaptations to prevailing conditions as well as short-term, rapid ecological fluctuations. Thus, teeth are frequently used to establish phylogenetic affinities among taxa. However, convergent evolution (homoplasy) may greatly distort actual phylogenetic relationships among closely related species.
Mammals show a heterodont dentition (different type of teeth depending on their function), unlike reptiles that most frequently show a homodont dentition. Homodontia is a derived condition inherited from the Permian Theraspsid reptiles. Another condition inherited from the reptiles is that mammals show two consecutive dentitions (diphyodont). The decidual (infant) teeth are replaced by the permanent (adult) ones. The diphyodont condition derives from the more primitive one in which teeth are continuously replaced as they wear away, as seen in sharks. The dental formula of the ancestral primate, described from the fossil record, most likely was 2143/3143, with 2 incisors (I), 1 canine (C), 4 premolars (P) and 3 molars (M) in the upper hemimaxilla, and 3 I, 1 C, 4 P, and 3 M in the lower hemimaxilla. Along the evolution of the different groups of mammals a significant reduction of teeth number may occur depending of the degree of dietary specialization. Humans show a dental formula of 2123/2123, which indicates a reduction of 2 upper premolars, 1 lower incisor, and 2 lower premolars, representing a shift from 42 to 32 teeth. Other mammal groups have much greatly reduced their teeth number. Within primates, only some lemurs have retained the four ancestral premolars, whereas most prosimians show 3 premolars (associated to a reduction of the snout), as also observed in the platyrrhines (New World monkeys). The catarrhines only retain P3 and P4, having lost the first and second premolars.
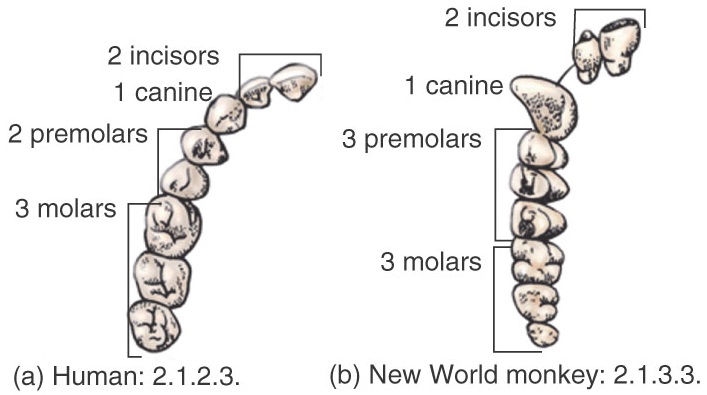
Dental formulae of humans and monkeys.
From an evolutionary perspective, the distinct shapes of teeth correspond to dietary adaptations to cutting and tearing of incisors and canines (anterior dentition) and to crushing and grinding of premolars and molars (posterior dentition). The conic morphology of the canine corresponds to the primitive condition of the homodont dentition of the reptiles. From this conic shape all tooth types evolved by developing additional cusps that extended tooth surface in response to a shift to a more omnivorous diet. The Tritubercular theory of tooth evolution assumes that homodont teeth evolved from a single-cusped conical tooth. As opposed, the Multitubercular theory suggests that the different types of teeth evolved through simplification of an ancestral tooth showing numerous cusps. However, the tritubercular theory most closely explains the great variability in dental shape observed in mammals.

Nomenclature of cusps in molars.
Dental cusps receive distinct names so we may specifically refer to them. The main cusp of a conical tooth is the protocone (Pr) in the upper dentition and the protoconid (Prd) in the lower dentition. From this ancestral condition, two cusps are added to provide a cutting edge: paraconid (Pa) and metaconid (Me) in the maxilla and paraconid (Pad) and metaconid (Med) in the mandible. Pa and Pad are mesially placed and Me and Med are distally placed. These three cusps lie in line providing a cutting border, as seen in the upper P4 (UP4) and lower M1 (LM1) of dogs. This linear dental shape is greatly efficient in carnivores for cutting flesh but is not suitable for crushing or grinding harder food items, such as plant foods or seed. In order to extend dental surfaces for grinding, the central cusps is displaced laterally, to the lingual side the Pr and to the buccal (vestibular) side the Prd. This produces a triangular shaped molar tooth called the tribosphenic molar. The upper and lower molar can extend distally with the addition of a talon and talonid (clearly separated from the trigon and trigonid that included the tree main cusps. The talon only includes one additional cusp, the hypocone (Hc), whereas the talonid may have up to three additional cusps: hypoconid (Hcd), hypoconulid (Hnd), entoconid (End). Initially, both the talon and talonid are clearly distinguished form the mesial trigon and trigonid, but their heights eventually meet. The majority of modern primates have a 4 cusped upper molars and a 6 cusped lower molars, but this condition greatly varies. In the catarrhines, the cercopithecoidea show a derived condition whereas the hominoidea retain, to a certain extent, the primitive one. In the hominoidea the upper molars have 4 cusps and show a distinct hypocone, and the lower molars lose the metacone in the trigonid and show 5 cusps in a specific Y5 configuration. The cercopithecoidea lose and additional cusp in the lower molars and both the upper and lower molars have a reorganized, 2 by 2 parallel cusp pattern with crests joining the buccal cusps with the lingual ones. This is a derived condition called bilophodontia, exclusive of the cercopithecoidea, whereas the hominoidea retain the ancestral bunodont configuration, lacking crests. The evolution of the talon and talonid serve to maximize dental occlusion and increase tooth-to-tooth contact while chewing food items. The evolution of the hypocone is supposed to be homoplastic (derived from convergent evolution) in different mammal groups, which is indicative of the significance of this anatomical trait as an adaptation to the consumption of tough diets.
The primitive condition of the hominoidea provides a distinctive anatomy of molar teeth that is very helpful to distinguish hominin fossil specimens from cercopitecoid ones in the East African record. The molar teeth of Australopithecus specimens retain the Y5 bunodont morphology that is clearly discernible from the bilophodont teeth of fossil baboons and Theropithecus. Hominin molar teeth show rounded cusps and a Y5 cusp pattern, whereas Papio and Theropithecus show 4 cusps arranged in parallel and joined by a bucco-lingual crest.
There are some evidences that the incisors and premolars also derived from the primitive tribosphenic morphology. Recently erupted incisors show the 3 so-called mammelons that suggest three cusps arranged in a row, though they rapidly wear away. In addition, some premolars may show a distally extended basing that deviates from the 2-cusped symmetric typical premolar shape. In fact, the canine is the only tooth that has retained the ancestral conical morphology. Modern humans may show some further simplification of dental cusp pattern. The LM2 shows a +4 cusp pattern (having lost an additional cusp), the UM2 may lose the hypocone, showing only 3 cusps (a frequent trait in the third molars), and in some cases premolars may reduce their lingual cusp or even duplicate it. The sectorial premolar is a primitive condition shared by many haplorrhini, but it lacks in humans and is present in the most ancient hominins. It is mostly present when the canine greatly extends the occlusal plane due to its length, and is associated to the presence of diatemae (empty spaces in the geniva) that allocate the oposed canine during occlusion. In this morphological condition the canines act as scisors by contacting laterally (scisor complex). This lateral contact between canines extends to the UC distal ridge that contacts with the proximal sector in the LP3. This sector (or ridge) is associated to the absence of the lingual cusp on the premolar (monocuspid premolar) and is especially evident in the papionini, extending to the premolar root.
The reduction of teeth number and dental complexity in the hominini is not highly significant. Humans show the same dental formula as all the catarrhines and only differ from them in having a reduced canine size, lack of diastemae and sectorial premolar, reduced +4 pattern in the lower second molars and even more reduced pattern in the third molars. This reduction in molar morphology in humans evidence a decreasing importance of bite forces for food processing. These dental evolutionary trend is acompanied by the reduction of muscle attachments and strength. In particular those related to the temporalis muscle that attaches to the coronoid process in the mandible, and to the masseter muscle that attaches to the malar inferior rim and the zigomatic arch. Non-human apes show strong temporal and masseter muscles, as well as great robusticity of the corresponding muscle attachments. Therefore, muscle development and cranial robusticity are significantly related to dental size, cusp pattern and dental enamel thickness.
d. The human brain
Brain expansion (a major adaptation in humans) started some 2 million years ago. However, the increase in brain size was the result of an increase in brain complexity and organization. The central nervous system is formed by the brain (cerebrum, cerebellum, brainstem) and the spinal cord. Additionally, the peripheral nervous system (cranial and spinal) is in charge of receiving and transmitting the sensitive impulses to the central nervous system and to the organs of the motor impulses. This vegetative neural system extends along the vertebral column and the thorax and abdomen, innervating the organs and muscles. This system is autonomous and can control the function of the major organs without the direct control of the central nervous system.

The main areas of the human brain (left hemisphere).
The brain (cerebrum) is formed by an external area (the cortex) showing multiple imbrications that extend its overall surface. The cerebrum is divided in various areas separated by sulcus or fissures. A sagital sulcus divides the brain into two hemispheres (left and right), each of which can be divided into lobes (frontal, parietal, occipital, temporal, limbic, and insula). Three main sulcus separate the lobes: 1) the Silvio sulcus (separating the temporal and parietal lobes), 2) the simian sulcus (perietal-occipital), and 3) the inter-hemispheric sulcus (separating the left and right hemispheres that are connected through the corpus callosum). Within the lobes, several cortical areas can be distinguished by their particular structure and/or function (named by Brodmann using numbers), providing a map of cortical functions of the brain. The frontal lobe is delimited posteriorly by the Rolando sulcus and its most anterior part is the prefrontal cortex (areas 9-15, 32, 43, 44). The frontal lobe also includes the motor areas (areas 4, 6, 8, 43, 44) and Broca’s area (44-45) that connects the visual and auditory areas with the motor cortex. The temporal lobe is located below the Silvio sulcus and includes the auditory perception areas (22, 41, 42), the visual cortex (areas 20, 21, 37), and the olfactory cortex (38). The limbic lobe is related to emotional and affective processes, learning and memory (areas 25, 33, 24, 31, 23, 30, 26, 29, 27, 35, 28, 34). The parietal lobe is located behind the frontal lobe and includes the sensitive areas 1, 2, 3, 5, and 7, which are in charge of functions related to spatial orientation and body perception. The occipital lobe includes the cortical areas 17, 18, and 19, as well as part of area 37. It is a mainly visual area related to perception of shape, stereoscopic depth, visual acuity, and colour, brightness and movement perception. Finally, the insula lobe is related to the perception and digestive functions.
The brain cortex (neo-cortex) consists of a thin layer of about 1 to 4 mm that contains around 100,000 million neurones forming different layers. The basal layer is formed by pyramidal neurones (over 75% of the cortex) that conduct the cortex responses through the thalamus. The hypothalamus, which includes the hypophysis gland, regulated the endocrine functions of the thyroid, suprarenal glans, pancreas and gonads. The hypothalamus also controls eating and drinking functions, body temperature, sexual responses and sleep. The cerebellum cortex makes adjustments to the motor responses that originate in other parts of the brain in relation to walking and equilibrium.
In most of the ancient vertebrates (reptiles and amphibians) the brain cortex is small and is mainly associated to olfactory functions, whereas in mammals the brain cortex is large and also includes tactile, visual and auditory functions. In primates the olfactory cortex is much reduced compared to other mammals. Sensory and motor brain areas are greatly connected in primates and their respective cortex areas are significantly expanded increasing cerebral convolutions.
i. Lateralization, language and handedness
In vertebrates, each brain hemisphere controls body functions in the opposite side of the body, although both hemispheres and functionally integrated through connections in the corpus callosum. Brain functions may be specialized in each brain hemisphere, causing brain lateralization and asymmetry. In humans there are areas of the cortex that are repeated in each hemisphere, connected by the corpus callosum, but other areas are unique in one hemisphere.
 In humans, language abilities are the result of interconnection of various cerebral areas in the brain controlling sensorial functions, stored information and motor functions. Language involves understanding and transmitting thoughts and feelings using sounds or gests that are understandable to others. This transmission requires rules that operate using the motor areas of the brain in relation to muscles controlling the lips, mouth, tongue, and larynx to produce and receive sounds. The brain cortical areas associated to language are placed in the dominant hemisphere, generally the left (90-95%). In about 70% of left-handed people the dominant hemisphere is also the left, and in the other 30% language functions are located in both hemispheres (50%) or in the right one (50%). Language associated areas, located in the left hemisphere, are either associative (Wernicke’s areas 41/42 and Brodmann’s area 39) or executive (Brodmann’s area 44/45). Wernicke’s area is located on the temporal lobe and is in charge of the perception of phonetic language, whereas area 39 is located on the parietal lobe and is related to perception of written language. Brodmann’s areas 44 and 45 correspond to Broca’s area, located on the frontal lobe and related to the motor functions associated to speech. The right hemisphere is involved in non-verbal expression, musical language, and emotions with facial expression and body gesticulation, whereas the left hemisphere is associated with verbal expression, the use of words and symbols, and mathematical language. The connection between both hemispheres allows the integration of those functions that are unique and specialized in each one of them.
In humans, language abilities are the result of interconnection of various cerebral areas in the brain controlling sensorial functions, stored information and motor functions. Language involves understanding and transmitting thoughts and feelings using sounds or gests that are understandable to others. This transmission requires rules that operate using the motor areas of the brain in relation to muscles controlling the lips, mouth, tongue, and larynx to produce and receive sounds. The brain cortical areas associated to language are placed in the dominant hemisphere, generally the left (90-95%). In about 70% of left-handed people the dominant hemisphere is also the left, and in the other 30% language functions are located in both hemispheres (50%) or in the right one (50%). Language associated areas, located in the left hemisphere, are either associative (Wernicke’s areas 41/42 and Brodmann’s area 39) or executive (Brodmann’s area 44/45). Wernicke’s area is located on the temporal lobe and is in charge of the perception of phonetic language, whereas area 39 is located on the parietal lobe and is related to perception of written language. Brodmann’s areas 44 and 45 correspond to Broca’s area, located on the frontal lobe and related to the motor functions associated to speech. The right hemisphere is involved in non-verbal expression, musical language, and emotions with facial expression and body gesticulation, whereas the left hemisphere is associated with verbal expression, the use of words and symbols, and mathematical language. The connection between both hemispheres allows the integration of those functions that are unique and specialized in each one of them.
ii. Variability and ontogeny
Modern humans show well-developed cortex areas of Wernicke and Broca, whereas within the mammals only the hominoidea show rudimentary Wernicke’s area. During development in humans, both hemispheres intervene in language communication until 4-5 years of age when the left hemisphere acquires greater eminence while the right one reduces its language attributions. Thus, the brain shows an initial undifferentiation until language is fixed as an specialized trait. Brain areas associated to language are highly genetically determined, although their development may be affected by environmental conditions. The right hemisphere may assume to a certain extent the functions associated to language skills until they are fixed at around that age. Brain lateralization may be completed in humans between 5 and 12 years of age. During this period, brain communication capabilities need to be stimulated to properly complete development.
Non-human pongids do not show a significant development of Broca’s area, whereas the chimpanzee shows a well-developed Wernicke’s area, which suggests that the anatomic lateralization associated to language capabilities was already present in the last common ancestor (LCA) between chimpanzees and humans some 8 million years ago (Ma). In fact, these areas are the last to complete neural development, which is consistent with the development seen in the great apes. In modern humans the great development of Wernicke’s area has displaced backwards the visual cortex in the occipital lobe. The brain cortex in humans is 3.2 times larger than that in non-human hominoidea. The area 39 (also known as angular gyrus) is especially associated to language, along with Wernicke’s area. They process the visual and tactile stimuli before constructing a syntactic response in Broca’s area that determines the action of the motor area that controls the muscles involved in the articulation of the phonetic responses.
iii. The limbic system
The limbic system is located at the base of the brain, in the most primitive region. It controls emotional response to basic instincts, such as fear, wrath, sex, smell, taste, thirst and hunger. Language can be used to relate two non-limbic impulses (non related to emotions), such as a sound and a word or a concept, whereas most mammals can only relate a non-limbic impuls with a limbic one and thus their behaviour is mostly oriented by emotions. The separation of the language areas and the limbic system allows humans to associate sounds (words) with concepts. The area of the brain cortex where this non-limbic association is made are Brodmann’s areas 39 and 40 that occupy a broad region in the parietal lobe besides Wernicke’s areas 41 and 42. These areas allow complex behaviours with little association to emotions, which may be the base of the symbolic behaviour in humans, chimpanzees, and perhaps also the orangutan.
e. Social behaviour of monkeys and apes
Social behaviours rarely preserve in the fossil record. The origin and evolution of our complex interactions and social structures needs to be inferred from both fossil specimens and the hard evidences of cultural practices. The cultures of present day hunter-gatherer populations is too recent in our evolutionary history to be representative of our most remote ancestors that most likely lacked our complex communication systems, mainly based on language. There are three main difficulties one needs to overcome when trying to infer the evolution of social behaviours in our lineage:
- Social models in extant primates are only indicative of the result of the evolution of behaviour and may not be representative of the social structures of past populations and species. In fact, behaviour is, to a great extent, a response to environmental conditions, and those prevailing now might not have been the same in the past. However, we may try to extrapolate some behaviours observed in living hominoidea to our ancestors and discuss wether they might be representative of their own social and behavioural organization.
- The source of behavioural knowledge in non-captive primates is scarce. Behaviour of captive animals might differ from that of non-captive ones and even the observation of their behaviour in liberty may modify the natural social patterns. Thus, building models of social structure behaviour , and mating systems in hominoidea primates may not be informative of their actual behaviour.
- Behaviour is often a subjective appreciation and, as humans, we tend to project our own culture and viewpoint to the interpretation of the behaviour of other species.
i. Primate models
The study of the evolution of behaviour may be interpreted from two main perspectives: phylogenetic and environmental. The phylogenetic perspective sustains that social behaviours such as agression, territoriality and cooperation have a great genetic component, as suggested by Sociobiology. Phylogenetic inheritance of behavioural patterns determines cultural patterns, which can not be modified despite environmental conditions change. For instance, Macaca nemestrina is able to recognize individuals of its own family group despite grown up separately. Similarly, Papio hamadryas maintains social behaviour even in captivity. These examples suggest that behaviour is highly inherited and phylogenetically dependent. On the other hand, the environmental perspective suggests that social structure and behaviour may change depending on environmental constraints. For instance, terrestrial species inhabit greatly variable environments and may shift from a large, multi-male structure to a small, single-male family group. In fact, sexual dimorphism and its associated difference in sex-related roles vary depending on the size of the occupied territory and the abundance of food sources. Behaviour may thus vary depending on evironmental conditions.
a. The baboon model
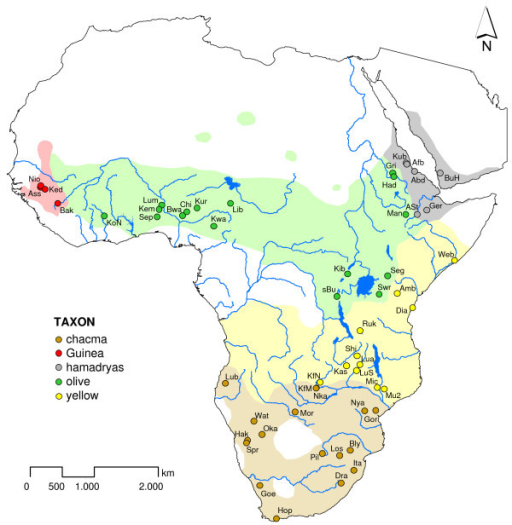
Geographical distribution of baboons.
Baboons are African and Arabian Old World monkeys belonging to the genus Papio, part of the subfamily Cercopithecinae. They include a minimum of five species that are some of the largest non-hominoid members of the primate order; only the mandrill and the drill are larger. Previously, the closely related gelada (genus Theropithecus) and the two species (mandrill and drill) of genus Mandrillus were grouped in the same genus, and are still often referred to as baboons in everyday speech. They range in size and weight of the baboons depending on species. The Guinea baboon is 50 cm (20 in) and weighs only 14 kg (31 lb), while the largest chacma baboon can be 120 cm (47 in) and weigh 40 kg (88 lb). Baboons (genus Papio) are terrestrial (ground dwelling) and are found in open savannah, open woodland and hills across Africa. Their diets are omnivorous, but mostly herbivorous, yet they eat insects and occasionally prey on fish, trout and salmon if available, shellfish, hares, birds, vervet monkeys, and small antelopes. They are foragers and are active at irregular times throughout the day and night. Baboons are distributed all over sub-Saharan Africa, excluding most parts of the west and central African rainforest, and have even colonized parts of the Arabian Peninsula. They form morphologically and geographically distinct populations but demonstrate no pre- or postzygotic reproductive isolation. Additionally, baboons are known to produce fertile hybrid offspring with geladas (Theropithecus gelada). The divergence between these sister genera is estimated at 3.5–4 ma. The five generally acknowledged baboon types are: a) the Guinea baboon (P. papio) from West Africa, b) the olive baboon (P. anubis) from the northern savannah belt, c) the hamadryas baboon (P. hamadryas) from north-east Africa and the south-western Arabian Peninsula, d) the yellow baboon (P. cynocephalus) from the East African coastal lowlands, Zambia and Angola, and d) the chacma baboon (P. ursinus) from southern Africa.
a)  b)
b)  c)
c)  d)
d)  e)
e)
Two cercopithecoidea, widely studied species are representative of this baboon model: Papio anubis (Olive baboon) and Macaca mulata (Rhesus monkey, not a baboon but sharing behavioural traits with the olive baboon, which reinforces the phylogenetic perspective of behavioural models). They form groups greatly variable in size, from 5 to almost 200, with average ranging from 30 to 50. During the day they search for food throughout the territory and at night they gather at a central area for resting at elevated rocks or tress. The territories of different groups may overlap and they tend to avoid contact when wandering in search for food. If they meet, for instance for drinking, they do not merge not exchange individuals. All groups show similar adult sex ratios (1 male / 2-3 females) and sexual maturation is earlier in females (3.5-5 years) than in males (7-10 years). Sexual dimorphism is also large (2 to 3 times in weight), which might be related to defense in front of predators (leopards, lions, hyenas), which results in a selection for great, dominant males. This may have had a great influence on the social structure of the baboons. The alpha male acts as a leader, defending the group and, to a certain extent, defining the mating patterns. Several males may occasionaly share the same status and collaborate in the defense of the group. Females also show a hierarchical organization and their status within the group may vary depending of their reproductive status or their association to the alpha male. The dominant males tend to occupy a central position in the group, while the youngs or less dominant ones occupy a perypheral position and may abandon the group and return. Females have a 35 days sexual cycle and show a sexual swelling during ovulation. In Papio anubis, mating takes place at the maximum of the estrogenic cycle and they separate. However, in Papio hamadryas male and female form durable bonds and the offspring may ‘inherit’ their status according to those relationships. Mother-child bonds are strong. The child hangs from the mother under the protection of the alpha male, until it climbs to the back. At the age of 6 months the child eats by itself until weaning at 11-15 months, when the ovulatory cycle of the female is resumed. Grooming (a reciprocal behaviour) is a major mechanism for social interaction. Females dedicate more time to grooming practices than male, either to groom on the dominant males, to other females or to their children. Grooming reinforces social interactions and relationships. Communication among baboons is mainly through vocalizations and visual signals. Vocalizations can denote attack, fear, sexor friendship, among others, and frequently acompany visual signals, such as facial expressions
The hamadryas baboon shows some behavioural differences from the olive baboon. It occupies more open areas and form large groups that can reach to 750 or more individuals, with an average of 136, as a result of the reunion of single-male units that split for food search. The single-male units consist of one or more females and their offspring, and some elder individuals. This basic unit remains together for life. The alpha male exercises a strong control on the group and mating is restricted to the group members. Some males may not have a female, mostly subadults, and if there is only one female in the group, mating is limitted by the estrogenic cycle. The single-male group may be formed either by 1) a subadult male kidnapping/adoption of a young female (6 to 12 months old), or 2) an adult male taking away the power and substituting the alpha male of a mating group.
b. The chimpanzee model
The social structure of the chimpanzee (Pan troglodytes) greatly differs from that of the baboons. They form much smaller groups that share a significantly more open structure in which the groups may split or combine alternatively, mostly depending on environmental conditions and food availability. This social structure is often called fusion-fision. Four main social groupings have been detected: 1) group of adult males without females or infants, with significant mobility; 2) groups of females with depending infants, without young females and sometimes with an elder male, generally more sedentary than the male groups 3) groups of both males and females, excluding young females, with intense sexual activity, mediated to a certain extent by hierarchy but also with promiscuous mating, and 4) groups of both males and females of any age, most frequently around a food source. As a whole, the chimpanzee shows a highly open social structure with great interactions among individuals and groups. Larger groups may be formed when food is available. Some differences exist between chimpanzees in the forest boundaries and those from savanna environments. In general, those from savannas tend to be smaller and clearly distinct, although their ranges might overlap, showing hierarchical structres among them. There seems to be a confrontation between adult males of the different groups, whereas females may shift from one group to the other more easily, likely to prevent endogamy. A different case is the pygmy chimpanzee (Pan paniscus). It is likely a derived behavioural model, greatly based on a matrilineal structure where hierarchy is defined by the females and competition and agonistic behaviours are prevented by sexual interactions. Despite these, the bonobos and the common chimpanzees share a common social organization based on ecological constraints for fusion and fision of the basic family units. Communication among chimpanzees are complex combinations of facial and body signals along with vocalization.
ii. The Plio-Pleistocene hominin model
The inferences from the behavioural models may greatly vary depending on the model used to extrapolate the behaviour of our most remote encestors:
- From the olive baboon models we may assume that the hominins, as primates living in savanna environments, would have formed large, mainly closed groups with a polyginic mating system with an alpha male showing significant sexual dimorphism (males in charge of the defense and females rearing the children) and with a well-stablished male hierarchy based on competition for females and for food.
- From the hamadryas baboon model we may infer that the overall social group might include stable units based on a single male with one or more females sharing strong bonds that would last beyond a single reproductive cycle.
- From the chimpanzee model in forested environments we may assume that the hominines from such closed environments would have shown open social structures, with great mobility of individuals. This open structure might have evolved to a more closed one with the shift from close forested environments to open savannas. In such open groups, males might collaborate for food quest, while females might collaborate in rearing the offspring and other purposes. Depending on food scarcity, the groups might split into basic family units. Hierarchy and sexual dominance might not have been significant in this social structure. Thus, sexual behaviour might have been highly promiscuous, although a basic monogamous unit might have coexisted within the group, especially for economic (food provisioning) or reproductive (child rearing) reasons depending on environmental conditions.
- From the chimpanzee model in more open environments we may asume that the hominines would have retained a semi-open structure limited by environmental constraints, with a significant antagonism between males of different groups, whereas females would move easily between groups to prevent consanguinity, a characteristic human trait.
Other social models might be considered as well for inferring hominin behaviours, such as the monogamous gibons. However, from a phylogenetic perspective, the models based on the chimpanzee are preferable for their evolutionary proximity to the hominines. A combination of inferences from the models described may also be appropriate.
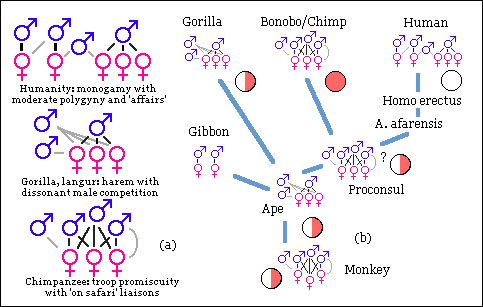
Hypothetical evolution of mating systems within the hominoidea.
From an evolutionary perspective, the monogamous social structure of the gibons seems derived from that of an ancestral ape, which might have had a polygynous mating system based on male competition. If this is the case, the polygyny of the gorilla’s might represent more closely the ancestral condition than that of the chimpanzees, with more open social structures. In contrast, the modern humans social structure would have derived from a promiscuous sexual patterns, as seen in chimpanzess, although with reduced levels of polygyny.
References
- Aboitinz F (2001) Size and complexity of the brain in human evolution. In: Tobias PV et al. (eds.) Humanity from African Naissance to coming millennia. Witwatersrand University Press, pp. 355-359.
- Aiello L, Dean C (1990) Human evolutionary anatomy. Academic Press, London-
- Annett M (1998) The brain and left-handedness. In: Jones S et al. (eds.) The Cambridge encyclopaedia of human evolution. Cambridge University Press.
- Campbell BG (1998) Human evolution. Aldine Publishing Co., Chicago.
- Chiarelli B (2005) Los primates actuales. In: Rebato E, Susanne C, Chiarelli B (eds.) La Antropología Biológica. Verbo Divino Ed. Estella, Navarra. pp- 207-220.
- Deacon TW (1992) The human brain. In: Jones S et al. (eds.) The Cambridge encyclopaedia of human evolution. Cambridge University Press.
- Eccles JC (1989) Evolución del cerebro. Ed. Labor, Barcelona.
- Fleagle JG (1999) Primate adaptation and evolution. Academic Press.
- Holovka S, Petitto A (2002) Left hemisphere cerebral specialization for babies while babbling. Scince 297, 115.
- Kardong KV (2006) Vertebrates. Comparative anatomy, Function and Evolution. McGraw Hill, New York.
- Kinzey WG (Ed.) (1987) The evolution of human behaviour: primate models. State University of New York Press.
- Le Gros Clarke WE (1971) The antecedents of man. Quadrangle Books, Chicago.
- Napier JR (1979) The antiquity of human walking. Scientific American, April 1967, pp. 50-60.
- Ojemann GA (1991) Cortical organization and language. The Journal of Neuroscience 11, 2281-2287.
- Quiat D, Reynolds V (1993) Primate bahaviour. Cambridge University Press.
- Reynolds V (1997) The evolution of human society. In Harryson et al. (Eds.) Human Biology. Oxford University Press.
- Sabater Pi J (1984) El chimpancé y los orígenes de la Cultura. Anthropos, Barcelona.
