a. Miocene ecology
The Miocene (23-5.3 Ma) constitutes the period in which the adaptive radiation of the Hominoidea primates takes place. 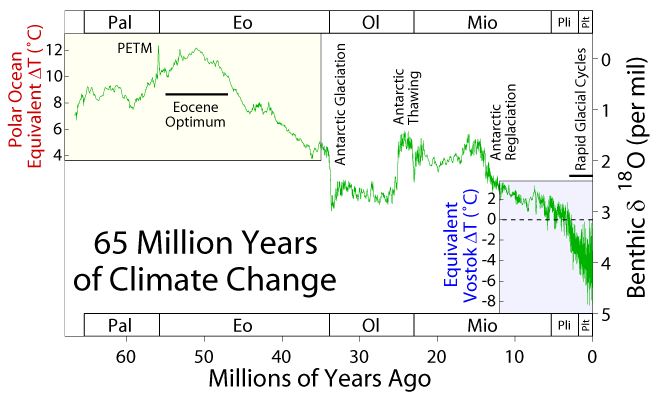 This was mainly due to the expansion of the tropical rain forest as a result of a progressive global warming. Overall temperatures significantly increased compared to those in the Oligocene. This climatic change was caused by various factors. Great tectonic activity took place forming the Alps, Himalaya, Pyrenees and the Atlas (North Africa) and Zagros (South Iran) mountain chains. At the Early Miocene (22-20 Ma) Africa was isolated from Eurasia. During the Middle Miocene (20-18 Ma) the Arabian Peninsula joined the Eurasian continent allowing the migration of the African primates into Eurasia.
This was mainly due to the expansion of the tropical rain forest as a result of a progressive global warming. Overall temperatures significantly increased compared to those in the Oligocene. This climatic change was caused by various factors. Great tectonic activity took place forming the Alps, Himalaya, Pyrenees and the Atlas (North Africa) and Zagros (South Iran) mountain chains. At the Early Miocene (22-20 Ma) Africa was isolated from Eurasia. During the Middle Miocene (20-18 Ma) the Arabian Peninsula joined the Eurasian continent allowing the migration of the African primates into Eurasia. 
At the beginning of the Terciary Era (∼65 Ma) subtropical and tropical forest dominated the Earth. Climatic conditions progressively decreased during the Eocene and Oligocene, favoring the diversification of the homeotherm mammals and birds, at the expense of the reptiles. At the begining of the Miocene, glacial periods are documented in the Antartida and Iceland, while open environments and savannahs expanded in equatorial areas and tropical forests abounded in Eurasia. By the end of the Miocene (around 7 Ma) global temperature had decreased again and most tropical environments in Eurasia disappeared again. During this period, the European hominoids went extinct. There is great controversy about the origin of the African Apes, either from hominoidea that survived through the climatic shift in Africa or from a group of hominoidea that migrated from Europe at the end of the Miocene. It is clear that the Eurasian hominoids migrated from Africa at around 20 Ma along with other mammals, such as the elefants. Hominoids ancestral to the hylobatids have been found at Ad Debtiya (Arabian Peninsula). Other fauna migrated at the same time into Africa from Eurasia, such as some suids, insectivores, carnivores (hyenas), bovids, cervids, giraffes, rhinoceros and rodents. These migratory movements were interrupted at around 16 Ma.
 The most ancient hominoidea remains, such as Proconsul and Kenyapithecus, were generalized quadrupeds lacking the brachiator adaptations that characterized extant hominoidea. The Pongidae and Hominidae lineages split between 16 and 14 Ma, represented by Sivapithecus in Asia and Dryopithecus in Europe. Well documented climatic changes at around 9.5 Ma caused the migration of hominoids to the East and South, disappearing from Europe at 7 Ma.
The most ancient hominoidea remains, such as Proconsul and Kenyapithecus, were generalized quadrupeds lacking the brachiator adaptations that characterized extant hominoidea. The Pongidae and Hominidae lineages split between 16 and 14 Ma, represented by Sivapithecus in Asia and Dryopithecus in Europe. Well documented climatic changes at around 9.5 Ma caused the migration of hominoids to the East and South, disappearing from Europe at 7 Ma.
b. European Miocene Apes
The genus Dryopithecus was the main European representative. It has been found in various sites aroung the Tethys sea in France, Austria, Germany, Spain and Hungary. The most ancient specimens of the genus seem to be those of D. fontani (12 Ma), D. laietanus and D. crusafonti (11-9 Ma), and D. brancoi (Rudabanya, Hungary, 11-10 Ma). Dryopithecus laietanus shows resemblances to the orangutan, gorilla and chimpanzee in the tempora bone, where it lacks the subarcuata fossa (present in all primates, including the hylobatids, but absent in D. laietanus as well as in the Great Apes. Its position as direct ancestor to the orangutan (as claimed by Moyà & Köeller (1995) is still debated, as Ankarapithecus has been suggested as ancestral to the Asiatic lineage (Kordos & Begun, 2002). Dryopithecus showed small vertebral bodies, indicative that their habitual locomotion was not bipedal. It also has longer arms than legs, indicative of a suspensory brachiator (Moyà & Köeller, 1996), a trait also present in chimpanzees and orangutans. This type of locomotion, that could be ancestral to bipedalism, might have appeared independently in the Asian and African hominoids. Dryopithecus was an arboreal primate that fed on fruits, similarly to the gorillas and chimpanzees. It was largely dristributed in Southern Europe where it survived during almost 4 Ma.
The species Oreopithecus bamboli dates to the end of the Miocene in Italy. It inhabited the forests in an island. It also lacks the fossa subarcuata, which indicates that it does not preceed the divergence of the hylobatids. It resembles Dryopithecus in limb proportion, the shape of the malar bone (flat, frontally oriented and with 3 foramina in the ascending ramus), and the shape of the zygomatic arch. However, Oreopithecus shows distinct traits in the coxal bone sugestive of bipedal locomotion, which would represent and independent acquisition of bipedal adaptations in relation to special ecological conditions. It also shows a short thumb (as in humans), an adaptation to precision grip. This would not be indicative of tool manufacturing, since Australopithecus also shows this trait in comparison to modern Apes, preceding in 2.5 Ma the use of lithic industries. This trait might have appeared as a response to dietary manipulation activities.
The species Pierolapithecus catalaunicus (13 Ma), that predates the split between the hylobatids and the Great Apes, shows the great variability of the Miocene hominoids in Europe.
c. Asian Miocene Apes
There is great consensus in considering Sivapithecus as a direct ancestor to the modern orangutan (Pongo). Sivapithecus indicus (Pakistan) and Sivapithecus (India) are the most representative species. They inhabited subtropical forests and would have been arboreal quadrupeds, lacking the brachiating specializations of the modern orangutan and showing a thiner dental enamel (indicative of soft fruit based diet) than that observed in modern orangutans. The facial morphology of Sivapithecus would be determinant in associating this genus to modern Pongo. However, other postcranial features are distinctive in Sivapithecus and may reflect some degree of convergent evolution in the adaptations to brachiation in the modern hominoids. The dating of Sivapithecus to between 12.5 and 7 Ma has been used to calibrate the molecular DNA hybridization clock. Gigantopithecus is a Late Miocene genus that may be descendant from S. indicus. Two species have been defined: G. bilaspurensis (Northern India), a giant species with massive mandibles and teeth, and G. blacki (a form dating to the initial Pleistocne), also a large specimen that might have coexisted with the first hominines in Eastern Asia around 2.0 Ma. Lufengpithecus lufengensis is a Late Miocene species from China, well documented but with highly fragmentary specimens.
d. Molecular clocks
 Mitochondrial DNA studies of modern Hominoidea show great molecular diversity among species, which suggests that the divergence time between species has been significant. Classical studies on DNA hybridization among hominoids have provided relevant information on the phylogenetic affinities of extant hominoids. Sibley & Alhquist (1984) provided evidence that chimpanzees and humans shared a greater proportion of DNA sequences that did gorillas and humans. The estimated molecular distances calculated based on DNA hybridization were then callibrated using the dating of Sivapithecus (estimated in 16-13 Ma in 1984). This analysis showed that the cercopitecoid primates split 33-27 Ma, the hylobatids did the same 22.4-18.2 Ma, the gorillas separated from our lineage 9.9-8 Ma, and humans and chimpanzees separated 7.7-6.3 Ma. Later, the dating of Sivapithecus was moved to 10.5-7 Ma, which suggest a maximum split between Pan and Homo of 5.5 Ma, 7 Ma for Gorilla and 2.0 for the split of the two chimpanzee species (P. troglodytes, P. paniscus). However, the short chronology for Sivapithecus would provide dates for the split of Homo-Pan not concordant with the fossil record of the hominines. Later studies have corroborated these classical studies, confirming the closer relationship between humans and chimpanzees, despite the greater morphological similarities of chimpanzees and gorillas. Different molecular clocks have been applied since then and different dates have been obtained, ranging from 4 to 7 Ma for the Homo-Pan lineage separation. This range of dates has significant consequences in the interpretation of early hominines and their anatomical adaptations, since a short chronology may consider some fossils as independent evolutionary paths, and a long chronology may include them as direct ancestors of our lineage. In fact, the fossil remains of Samburupithecus kiptalami (a 9.5 Ma possible ancestor of the gorilla), Orrorin tugenensis (a 6 Ma biped) and Ardipithecus kadaba (5.2 Ma) do not conform with such molecular chronology. A new estimation by Arnason et al. (2000) has suggested a Homo-Pan split at 13-10.5 Ma, which might fit better with the fossil record.
Mitochondrial DNA studies of modern Hominoidea show great molecular diversity among species, which suggests that the divergence time between species has been significant. Classical studies on DNA hybridization among hominoids have provided relevant information on the phylogenetic affinities of extant hominoids. Sibley & Alhquist (1984) provided evidence that chimpanzees and humans shared a greater proportion of DNA sequences that did gorillas and humans. The estimated molecular distances calculated based on DNA hybridization were then callibrated using the dating of Sivapithecus (estimated in 16-13 Ma in 1984). This analysis showed that the cercopitecoid primates split 33-27 Ma, the hylobatids did the same 22.4-18.2 Ma, the gorillas separated from our lineage 9.9-8 Ma, and humans and chimpanzees separated 7.7-6.3 Ma. Later, the dating of Sivapithecus was moved to 10.5-7 Ma, which suggest a maximum split between Pan and Homo of 5.5 Ma, 7 Ma for Gorilla and 2.0 for the split of the two chimpanzee species (P. troglodytes, P. paniscus). However, the short chronology for Sivapithecus would provide dates for the split of Homo-Pan not concordant with the fossil record of the hominines. Later studies have corroborated these classical studies, confirming the closer relationship between humans and chimpanzees, despite the greater morphological similarities of chimpanzees and gorillas. Different molecular clocks have been applied since then and different dates have been obtained, ranging from 4 to 7 Ma for the Homo-Pan lineage separation. This range of dates has significant consequences in the interpretation of early hominines and their anatomical adaptations, since a short chronology may consider some fossils as independent evolutionary paths, and a long chronology may include them as direct ancestors of our lineage. In fact, the fossil remains of Samburupithecus kiptalami (a 9.5 Ma possible ancestor of the gorilla), Orrorin tugenensis (a 6 Ma biped) and Ardipithecus kadaba (5.2 Ma) do not conform with such molecular chronology. A new estimation by Arnason et al. (2000) has suggested a Homo-Pan split at 13-10.5 Ma, which might fit better with the fossil record.
More recently Ebersberger et al. (2007) have reevaluated the molecular divergence among the hominoidea. Their results sugest that the human genome is a mosaic with respect to its evolutionary history. Based on a phylogenetic analysis of 23,210 DNA sequence alignments from human, chimpanzee, gorilla, orangutan, and rhesus monkeys, they presented a map of human genetic ancestry. For about 23% of our genome, we share no immediate genetic ancestry with our closest living relative, the chimpanzee. This encompasses genes and exons to the same extent as intergenic regions. About 1/3 of our genes started to evolve as human-specific lineages before the differentiation of human, chimps and gorillas took place. This explains recurrent findings of very old human-specific morphological traits in the fossil record, which predate the recent emergence of the human species about 5-6 Ma. The sorting of such ancestral phenotypic polymorphisms in subsequent speciation events provides a parsimonious explanation why evolutionary derived characteristics are shared among species that are not each other’s closest relatives. These results suggest that different sequences of the genomes of humans, chimps and gorillas have evolved at different rates and that convergence and divergence of both molecular and morphological traits have taken plave in the evolution of the hominoids.
e. The origin of the hominines
The Great Rift in Easter Africa, formed between 20 and 4 Ma, can be considered the cradle of the hominines, our lineage after the split of the chimpanzees. The Rift Valley separated the evolutionary history of the hominini in the East from that of the panini and gorillini in the West (McBrearty & Jablonski, 2005). This view was stated for the first time as the East Side Story by the French paleontologist Yves Coppens. The Rift valley extends more that 8,000 Km from the Dead Sea in the Near East to the Zambeze river in Zambia and Mozambique, with a bifurcation at both sides of Lake Victoria. The Oriental Rift, that crosses Ethiopia (Ethiopian Rift), including the Afar region, Kenya and Tanzania (Gregory Rift), and Malawi (Malawi Rift), holds the most abundant record of fossil hominines. In South Ethiopia and North of Kenya the valley was excavated by the Omo river. In this area the paleontological record of the hominines extends from more that 4 Ma to present days. At Olduvai, in Tanzania, a well documented record of hominini dates back to 2 Ma. The orogenic and volcanic activities at the Rift Valley had a great impact in hominin evolution, especially at around 8 and 4 Ma. The geologic changes associated to the formation of the Rift Valley resulted in a climatic separation of the East, more dry, and West, more humid, sides of the Rift. The endorreic concavities between the elevated lands (over 2,500 m at present) in the Rift were especially adequate for life, and were rapidly occupied by different types of plants and animals. Alternative processes of sedimentation and erosion have resulted in the exposition of layer deposits that include fossil specimens of our direct ancestors, the hominines.
The progressive climatic shift towards more arid and colder conditions during the Plio-Pleistocene forced new adaptation strategies that gradually separated non-human apes in the West from the human Apes in the East. This geographic disruption, along with the climatic shift, underly the origin and evolutionary process of our most direct ancestors. There is great debate, however, about how this process took place and if it was a unique event or included several instances before the hominini could evolve from a common ancestor with the chimpanzee.
References
- Andrews P (1987) Cladistic relationships of extant and fossil hominoids. J Hum Evol 16, 101-118.
- Begun (2001) African and Eurasian Miocene hominoids and the origins of the Hominidae. In: Phylogeny of the Neogene. Cambridge University Press.
- Begun DR, Nargolwaya MC (2004) Late Miocene Hominid Biogeography. Evol Anthropol 13, 234-238.
- Britten RJ (2002) Divergence between samples of chimpanzee and human DNA sequences is 5%, counting indels. PNAS 99, 13633-13635.
- Ebersberger I, Galgozy P, et al. (2006) Multiplex amplification of the mammoth mitochondrial genome and the evolution of Elephantidae. Nature 439(7077), 724-727.
- Fleagle JC (1999) Primate adaptation and evolution. Academic Press Inc, London.
- Jobling MA, Hurles ME, Tyler-Smith E (2004) Human evolutionary genetics. Taylor & Francis, New York.
- Klein J & Takahata N (2002) Where do we come from? The molecular evidence for human descent. Springer. New York.
- Kordos L, Begun DR (2002) Rudabánya: a Late Miocene subtropical swamp deposit evidence of the origin of the African Apes and humans. Evol Anthropol 11, 45-57.
- Martin RD (1990) Primate origins and evolution. Princeton University Press.
- McBrearty S, Jablonski NG (2005) First fossil chimpanzee. Nature 437, 105-108.
- Moyà-Solà S, Köehler M (1993) Recent discoveries of Dryopithecus shed new light on evolution of great Apes. Nature 365, 543-545.
- Moyà-Solà S, Köehler M (1993) New partial cranium of Dryopithecus Lartet, 1863 (Hominoidea, Primates) from the Upper Miocene of Can Llobateres, Barcelona, Spain. J Hum Evol 29, 101-139.
- Moyà-Solà S, Köehler M (1993) A Dryopithecus skeleton and the origin of the great-ape locomotion. Nature 379, 156-159.
- Navarro AN & Barton NH (2003) Chromosomal speciation and molecular divergence, Accelerated evolution in rearranged chromosomes. Science 300, 321-324.
- Ruvolo M et al (1994) Gene trees and Hominoid phylogeny. PNAS 91, 8900- 8904.
- Ruvolo M (1996) A new approach to studying modern human origins: hypothesis testing with coalescence time distributions. Mol Phyl & Evol 5, 202-219.
- Sarich V & Wilson A (1967) Inmunological time scale for hominid evolution. Science 158, 1200-1203.
- Seuánez HN (1987) The chromosomes of man: evolutionary considerations. Springer-Verlag.
- Sibley C, Alhquist J (1984) Thy phylogeny of the hominoid primates as induced by DNA-DNA hybridation. J Mol Evol 20, 2-15.
