a. Humans in Nature
Modern humans (Homo sapiens) share derived, specialized traits that have evolved as a result of long-term selective pressures acting under fluctuating climatic conditions in Africa. Among these, brain encephalization, enlarged growth and maturation (gestation, infancy, reproduction, life expectancy), and an articulated language, have evolved along the last 2.5 million years (Ma), since the emergence of the genus Homo, and especially so since the emergence of modern human populations about 200.000 years ago.
The increased encephalization and exploitation of a cultural niche, based on parental care and progressive acquisition of linguistic skills, have been crucial to human survival through time. Morphological evolution shows reduced significance with regard to social and technological adaptations (fabrications and use of tools) that, to a certain extent, have replaced the requirement for muscles strength or dental specializations (Fig. 1).
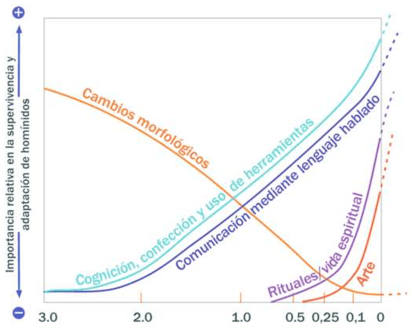
Figure 1. Relative significance of anatomical and cultural traits in human evolution (Tobias, 1997).
However, morphological adaptations have significantly shaped our bodies in relation to climatic and ecological conditions even at very recent times and are still doing so in extant populations.The long life expectancy and retarded maturation in humans, compared to other apes, provide time for brain maturation after birth and for social learning and integration. These biological and cultural adaptations likely evolved in Homo erectus, a direct ancestor to Homo sapiens that lived in Africa between 1.8 and 0.3 Ma, but great debate remains concerning the origin of the genus Homo about 2.5 Ma and if such derived traits were already present at that time. There is a general consensus on the attribution of this genus name to the fossil remains of Homo habilis despite their brain adaptations and ecological behavior greatly vary (see chapter 7). Humans cannot be studied apart from other mammalian species, no matter how distinct our anatomy is or if our culture is more sophisticated than that of other animals. Paleoanthropology is the scientific discipline that studies the origin, evolution and dispersal, both in time and space, of the hominin lineage.
We belong to the Homo sapiens species. The Linnaean taxonomic system, defined by Carolus Linnaeus (1707-1778) (Systema Naturae 1735), establishes that all species are referred to with a double (dichotomic) name including the genus name (Homo) and the species attribute that should always include the genus name (Homo sapiens). In case that several distinct populations can be distinguished within a species, a subspecies attribute may be added (Homo sapiens sapiens). The species names and attributes should always be written in italics or underlined (Homo sapiens) and the genus can be shortened (H. sapiens), as well as the species attribute when the subspecies name is used (H. s. sapiens). Linnaeus defined two human species: Homo sylvestris and Homo sapiens, and within the latter he described six subspecies (H. s. ferus, H. s. monstruosus, H. s. afer, H. s. europaeus, H. s. asiaticus and H. s. americanus). Despite this classification was wrong, Linnaeus defined a precise way to classify organisms into a hierarquical way in which closely related species had to be grouped into the same taxonomic clade.
Taxonomy is the set of rules that assigns a species name to an organism and classifies it into taxonomic groups according to the similarities of its anatomical and genetic traits with other species. In fact, the taxonomy of a group of organisms should reflect their phylogeny, their ancestor-descendant relationships. Taxonomic groups should be monophyletic, which means that they have to share a unique common ancestor. If this requirement is not met, the group is said to be polyphyletic. Humans have traditionally been classified into a distinct family (F. Hominidae), separated from the chimpanzees (Pan troglodytes), gorillas (Gorilla gorilla) and orangutans (Pongo pygmaeus) that were classified into a separate family (F. Pongidae). However, chimpanzees and humans share a unique common ancestor closer in time than those shared with the other hominoidea (Sp. F. Hominoidea), formerly called pongids. Therefore, classifying humans alone into a distinct taxonomic clade does not reflect the actual phylogeny of the group. Humans, chimpanzees, gorillas and orangutans should be all classified into the same family. Thus, the term hominids no longer refers exclusively to the human ancestors, but to the ancestors of all the great apes, excluding the gibbons, referred to as lesser apes. Since we belong to the hominini tribe, our ancestors should be referred to as the hominines, instead of the hominids, which is a broader, more inclusive category (see section 1f).
b. Adaptation and adaptability
Encephalization seems to be the main adaptive response to environmental pressures in humans. However, enlarged brains forced longer infant maturation. Adaptive constraints in relation to the maximum size of the birth canal and bipedalism (see section 2b) conditioned human life history through the modification of growth and development. Encephalization improve adaptation, in terms of differential survivorship and reproduction. Nevertheless, if adaptation involves greater specialization, adaptability is likely to decrease with adaptation. But adaptation and specialization are not synonymous. An efficient adaptation may result in a greater specialization, such as the bilophodont dentition of the cercopitecoid primates adapted to a folivorous diet, as well as in a generalized trait, such as the bunodont dentition of the apes, more appropriate for a more omnivorous diet. Adaptability refers to the capacity to adapt through cultural behaviors to fluctuating environmental conditions.

Figure 2. The morphological diversity of human populations (Mützel 1904).
Under certain circumstances biological adaptations would be more efficient in the long term than cultural adaptations. Genetic and morphological diversity (Fig. 2) are crucial to biological adaptation. However, human adaptability is greatly based on brain encephalization, which provides a cultural capacity to temperate or even modify the environmental constraints. The human brain is a highly specialized adaptation that allows a huge adaptability through culture, non-genetic determined behaviors. All organisms have the biological capacity of acclimatization to environmental shifts, at least to a certain extent, but cultural behaviors greatly increase such capacity.
Despite humans share highly sophisticated and complex technological traditions, they are not the sole animal species that have a distinct culture. Chimpanzees also show cultural traditions, tough less developed compared to those of humans. However, brain encephalization is no unique to humans. Dolphins, chimpanzees, some monkeys and even some mice species show brain sizes larger than expected in relation to body size (Fig. 3).
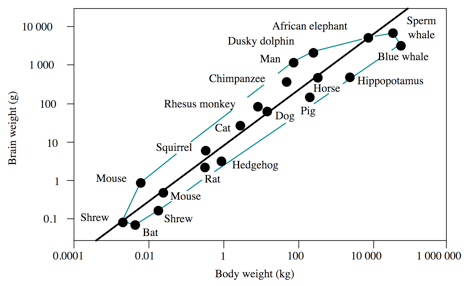
Figure 3. Brain weight relative to body weight in mammals.
Among mammals, dolphins and chimpanzees share a high encephalization quotient; modern humans have a distinct level of brain complexity and volume that allow them to modify the environment, so biological adaptation is replaced by cultural adaptation, which requires to efficiently transmitting culture through an articulated language. Therefore, the cultural evolution of humans require complex biological specialization, in particular those related to oral communication, growth and development, and reproductive strategies. Among primates, humans have an encephalization quotient (EQ) of 2.96; they have a brain volume respect to body weight almost 3 times larger than the average primate, followed by Saimiri (2.13), Miopithecus (2.00), and Cebus (1.98), whereas the chimpanzee (Pan troglodytes) has an EQ of 1.16. Brain size alone, thus, is not a reliable indicator of cultural complexity and adaptability.
The evolution of the hominin brain involves both size and reorganizational changes. The earliest australopithecines (A. afarensis) show a reduction of the primary visual cortex (area 17 of the brain) and a lunate sulcus in a posterior position by 3.5-3 Ma, which suggest that the parietal association cortex had increased compared to other apes. However, this species only shows a slight and allometric brain size increase to 400-450 cc. Cerebral asymmetry is also present but this is more evident in early Homo that in Australopithecus. Homo habilis shows a significant increase in brain size to 750 cc at 2 Ma, as well as reorganization of the frontal lobe and an increase in cerebral asymmetry. These modifications are coincident with stone tools and evidence of hunting and scavenging. The emergence of H. erectus involved a modest increase in brain size to 700-950 cc and increase in cerebral asymmetry. Brain evolution to 1,400 cc in H. sapiens involved both allometric and non-allometric changes, including refinement of cortical organization. Thus, the evolution of the hominin brain included size changes, reorganization of its structure and increase in asymmetry.
c. Evolution and heterochrony

Figure 4. The term humans includes all the hominin species included in the genus Homo.
Humans (Fig. 4) can be defined by various, very distinct traits that are unique to our species. Such anatomical derived characteristics are frequently referred to as autapomorphies. Our most distinct autapomorphies compared to the other hominoidea primates are related to our bipedal locomotion, large brains, reduced dental and facial traits, and enlarged life history and life expectancy.
Humans (Homo sapiens) generally inhabit open, non-forested environments, but at present are ubiquitous (occupy all possible habitats) by migration; morphologically we are non-specialized primates, although we have a highly specialized bipedal locomotion and a significantly large and complex brain that resulted in many behavioral and biological adaptations, such as slow sexual maturation, concealed (not visible) ovulation, complex social organization, high dependence on technology, food storage, hunter-gatherer life-style, or the use of articulated language for communication. All these derived anatomical traits have been shaped by natural selection to allow us to cope with specific needs during our evolutionary history.
Humans show the largest gestation period among primates. Despite this, the human newborn is highly dependent and cannot walk until about 9-13 months of age and brain reorganization and growth are very fast until 4 years of age. According to the adult brain size and compared to other primates, the human gestation should extend till 18 months of age. Thus, human birth occurs much earlier than brain development attains the same maturation as that seen in the chimpanzee’s newborn. A profound ontogenetic reorganization took place to allow premature babies and extent infancy to assure growth and development after birth, which increased infant dependence and parental care. Such a delayed maturation along evolution of a species is frequently referred to as neoteny, a type of heterochrony (Fig. 5), which can be defined as a shift in growth or development rates along the evolution of the lineage. Heterochrony can be either paidomorphic (retarded development) of peramorphic (extended development).

Figure 5. Evolutionary processes acting on ontogenetic development.
Paidomorphosis consists of the retention of juvenile traits of the ancestor in the later phases of the development of the descendant. There are three types of paidomorphosis: progenesis (premature end of somatic growth), neoteny (slow somatic growth), and postdisplacement (late somatic growth). Peramorphosis consists of an extension of the somatic growth beyond the adult condition in the ancestor. Three types of peramorphosis can be distinguished: hipermorphosis (late end of development), acceleration (quick rate of development), and predisplacement (early start of development). Many anatomical traits in human populations are considered neotenic since result from a slow somatic growth and maturation that results in an adult that retains the juvenile characteristics of its ancestor. Modern humans show rounded and gracile frontal bones lacking a robust supraorbital torus, reduced ears, short and rounded noses, and flat and vertical faces. All these anatomical traits are similar to those present in the infants of our ancestors and can therefore be considered neotenic.
Such evolutionary trend can be explained by natural selection affecting the so-called Hox (homeotic) genes. These are regulatory genes that act upon many structural genes responsible for the anatomical characteristics. Hox genes may delay or accelerate the activation of structural genes, thus affecting the development of anatomical structures. Mutations affecting Hox genes may significantly change the evolutionary history of a species, such as humans, and are crucial to understand ontogenetic processes and how these may be modified through evolution. On the light of the Hox genes, the old debate between Punctuated Equilibrium and the Neo-Darwinian Synthetic evolutionary processes can be dismissed. The tempo and mode of evolution are greatly affected by this type of genes and mutation rates need not to increase to explain many heterochonies in evolution.
d. Synthetic Theory and Cladistics
Evolution is a process by which populations acquire, modify and transmit their features (both anatomical and physiological) through time giving rise to new species with adaptations to varying environmental conditions. Mutation is the sole evolutionary factor that can generate new genetic variability and, hence, new anatomical adaptations, and the other evolutionary processes (natural selection, migration, genetic flow, genetic drift) modify the frequency of the morphological variation in the population. We now know that the theory of the inheritance of acquired traits is wrong since only mutations affecting the genetic information stored in the germinal cells can be transmitted from one generation to the next. The combination of the laws of inheritance developed by G. Mendel with the Natural Selection hypothesis by C. Darwin set the foundations of modern evolutionary science. The Synthetic Theory of evolution explains how evolutionary factors are responsible for the anatomical adaptation and evolution of populations.
From a population genetics perspective, a species is a natural group of individuals, reproductively isolated from others, that can interbreed and produce fertile descendants. The breeding factor can be easily confirmed in extant species but it may be more difficult to determine in extinct species. The concept of evolutionary species tries to solve this uncertainty by defining a lineage as a sequence of ancestor-descendant populations that evolves separately from other lineages, with unique evolutionary trends and specificities. Humans, as most mammal species, constitute a polytypic species that shows great geographic variation. With the introduction of the Cladistic theory, a species was considered the smallest diagnosable cluster of individuals within which an ancestor-descendant relationship can be defined. This definition lies on the characterization of traits that are primitive (plesiomorphic) or derived (apomorphic). Thus, phylogenetic affinities between species are set based on morphological similarities after the anatomical traits are classified and their state (presence or absence) and significance (weight) are clarified. The definition of the state of a trait in a polytypic species is frequently problematic since great variability may exist in the expression of the character. The distinct approaches defined by the Synthetic Evolutionary and the Cladistic views have given raise to two distinct classification procedures: Evolutionary Systematics and Phylogenetic Systematics. When studying human evolution the evolutionary view cannot be always applied, since the great variability of populations make it difficult to determine the boundaries of species in the absence of interbreeding information. Furthermore, the cladistic or phylogenetic view tends to disregard the polytypia of hominin species.

Figure 6. Gradistic and cladistic terminology for anatomical traits within a phylogeny.
Nevertheless, the cladistic approach provides a terminological framework and a methodological procedure to classify both geographic and temporal (chronospecies) populations in terms of an ancestor-descendant relationship. There is a clear equivalence between the evolutionary and cladistic terminology that can be used as synonymous (Fig. 6). Homologous traits are those inherited by two taxa (or OTUs, operational taxonomic unit) from a common ancestor and have been passed from generation to generation. On the other hand, analogous traits result from independent evolutionary processes and despite they may resemble in different OTUs, the taxa share a common ancestor that predates the independent origin of that trait in both lineages. Therefore, analogies do not indicate any type of phylogenetic relationship among taxa, whereas homologies are informative of species diversification. Primitive traits are those that have not significantly changed through time and derived traits are those that have suffered some degree of evolutionary change as a result of directional selection and adaptation processes. Darwinian evolution assumes that anatomical resemblance between taxa is indicative of a recent common ancestor from which they inherited the homologous traits. However, the speciation process might have modified such traits along the course of their particular evolutionary history. Since they are homologous traits, they were present in their last common ancestor (LCA). Derived traits differ in the descendants and are indicative of morphological diversification as a response to distinct evolutionary processes.
Cladism is a method for classifying and analyzing anatomical traits to establish ancestor-descendant relationships among taxa, in a similar way as Darwinian evolution does by classifying traits into primitive o derived. Most frequently, anatomical traits have been the basis for the taxonomic classification of species, although genetic information of extant species is of great relevance for this purpose. Cladistic researchers have developed a useful terminology (Fig. 6) for specifying if a trait is primitive or derived. For this purpose, Cladism uses the terms plesiomorphic (a primitive trait present in the LCA of a phylogeny) and apomorphic (a derived trait not present in the LCA but in a recent ancestor of a subclade within the phylogeny). If a plesiomorphy is shared by several extant OTUs it is called a simplesiomorphy. A shared apomorphy is called a sinapomorphy, and a trait that is unique to a single species or OTU is called an autapomorphy. Finally, analogous traits that result from convergent evolution are called homoplasies. These terms constitute the state of an anatomical trait, which along its weight are significant for the interpretation of the phylogenetic affinities among species. However, researcher subjectively interprets the state and weight of a morphological character and in many instances it is really difficult to make a proper decision. For instance, the uniqueness of human brain size and the complexity of our specialized locomotor bipedalism have been interpreted as relevant autapomorphies of our genus that justified excluding us from the Pongidae family, classifying our species alone into the Hominidae family. In this way, a taxonomic dichotomy between pongids and hominids was set (Fig. 7) despite it does not reflect the actual phylogenetic relationships between humans and the other apes. Molecular analyses of both mitochondrial and nuclear DNA have demonstrated that humans and chimpanzees share a close LCA about 5.5 Ma, whereas the common ancestors of humans with the other apes are much older.

Figure 7. Cladistic phylogenetic tree emphasizing the derived traits of humans, which results in a wrong taxonimic classification of humans alone into the Hominidae family.
In the phylogenetic tree in figure 7, the pongidae family constitutes a polyphyletic group, since it is based on overall anatomical similarities and does not include humans despite we share significant (mostly molecular) traits with them, and particularly with the chimpanzee. A phylogenetic tree should be monophyletic and the taxonomic classification has to reflect the actual ancestor-descendant affinities among species. The great morphological diversification of humans with respect to the other hominoids may reflect mutation and selection acting on regulatory genes determining ontogenetic processes and, thus, such significant degree of divergence should not be considered for phylogenetic classification.
e. Phylogenetic processes
New species appear through the interaction of mutation, selection and genetic drift along a lineage in which reproductively isolated species show an ancestor-descendant relationship. The fossils are representatives of those intermediate species that existed in the past. Fossil species are known as chronospecies. Reproductive interbreeding between chronospecies cannot be determined and, thus, the boundaries between taxa are defined based on anatomical traits. Along the evolution of lineages several speciation processes can be distinguished. Anagenetic speciation occurs along a lineage through time and, thus, result in the origin of chronospecies. Brain evolution in the genus Homo is a clear example of anagenesis since H. habilis, H. erectus, H. heidelbergensis, and H. sapiens can be considered distinct chronospecies with increasingly higher brain volume from 550 to 1400 cc. It is also a good example of gradual evolutionary processes. Within anagenetic lineages the rate of evolution can significantly slow down (stasigenesis) producing fewer anatomical diversity and a reduced number of descendant taxa. In contrast, cladogenetic speciation results in a branching process along a lineage that originates two distinct descendant clades. Cladogenesis is the result of significant changes in environmental conditions or the emergence of geographic barriers that separate populations. Allopatric speciation involves geographic separation of populations and frequently results in faster speciation processes, whereas sympatric speciation (speciation without geographic barriers between populations) is likely to be much slower and less frequent. However, reproductive barriers, other that geographic separation, are also required in sympatric speciation, though these could be behavioral. Reproductive barriers may not result in speciation. Parallel evolution in distinct taxa would minimize evolutionary diversification, while convergent evolution would tend to increase anatomical similarities between taxa, which Cladism calls homoplasies. These convergent traits not only are uninformative of actual phylogenetic relationships, but also tend to increase discrepancies between morphological and genetic phylogenies. Various anatomical traits in the apes are subjected to great debate concerning the type of evolution they have gone through: convergent, stabilizing, or diversifying. Even brain size has been sometimes considered a convergent trait between modern human populations and Neandertals.
f. Hominid Systematics

Figure 8. Taxonomic classification of Primates, including the Hominini, in agreement with molecular data.
The taxonomic classification of species, independently of the classification method used (gradualist or cladistic), should reflect the actual phylogeny of the species considered. Systematics is the study of the similarities among organisms and their phylogenetic relationships. It is clear now that establishing anatomical similarities between taxa requires defining the state of a character (whether it is a primitive or derived trait) so it may be used for phylogenetic inference. This is not always an easy task. As a general rule, evolution produces natural groups and the evolutionary factors are responsible for discontinuities among taxa. Therefore, the taxonomic classification should be exclusively based on genealogy (ancestor-descendant relationship) resulting in a phylogenetic classification (mostly based on molecular data). However, taxonomy should also reflect, at least to a certain extent, the anatomical similarities and differences among taxa, producing a phenetic classification (based mainly on anatomical data). The phenetic classification of organisms tends to classify organisms into distinct grades that include species that share similar anatomical adaptations. However, the phylogenetic classification cluster species is based on clades that reflect actual phylogenetic affinities among species. Grades and clades generally coincide but convergent evolution (homoplasies) tends to obscure the phenetic classification.
Humans form a distinct grade since they share relevant adaptations, such as brain evolution and ontogenetic development, and they also form a clear clade since can be differentiated based on their anatomical resemblances. However, out taxonomic classification within the primates has been the subject of great debate, mostly because of the difficulties in assuming that the phylogenetic classification should be reflected in the taxonomy of the group despite a phenetic classification and homoplasies may be misleading in doing so. Molecular analyses have undoubtedly demonstrated that the chimpanzee and humans share a common ancestor that is closer in time to those shared by the chimpanzee and the other great apes. Thus, Homo and Pan should be grouped into a common taxonomic unit (subfamily Homininae). Then, the Homo-Pan clade shares a closer common ancestor wit Gorilla than with any other ape and these later cluster with the orangutan (Pongo). Therefore, the greater apes should be classified together (family Hominidae), closely related to the lesser apes (family Hylobatidae). These two families may then be grouped into the Hominoidea superfamily (Fig. 8). In this way, the term hominid no longer refers to our species and exclusive ancestors, but to all the great apes and their ancestors, including humans. If we want to refer to our lineage since we diverged from the chimpanzee lineage, we should use the term Hominini. Hominini appeared around 6 to 5 Ma in Africa. We will dedicate the following chapters to understand how the hominini originated and what evolutionary trends characterize our lineage.
References
- Tobias PV (1997) Evolution of the brain size, morphological restructuring and longevity in early hominids. In: Principles of neural aging (Dani SU, Hori A, Walter GF Eds.). Elsevier pp. 153-174.
- Mützel G (1904) Asiatiska folk, Nordisk familjebok vol.2.
