La extinción masiva de la transición Cretácico/Terciario (K/T), hace unos 65 millones de años, redujo en torno al 11% la diversidad taxonómica a nivel de Familias. En particular, desaparecieron los dinosaurios, lo que posibilitó la radiación evolutiva de los mamíferos. Esta 5ª extinción se produjo muy probablemente por el cambio climático causado por la colisión de un asteroide (Sci Am 1990, Oct pp. 78-85) en la península de Yucatán que causó el cráter Chicxulub que aún es posible detectar en los fondos marinos y la costa mexicana.
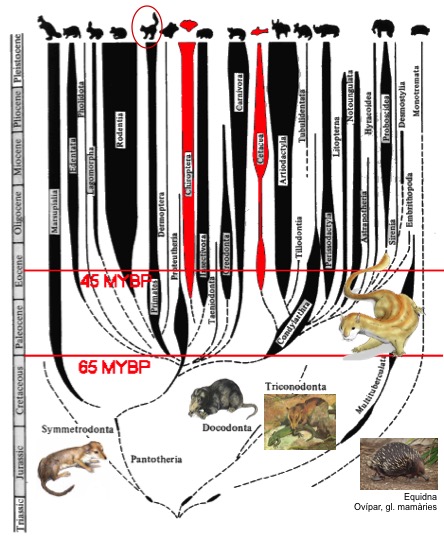 La respuesta biótica a una extinción masiva como la que causó la desaparición de los dinosaurios es la radiación adaptativa. En la transición K/T los grandes beneficiados fueron los mamíferos que experimentaron un rápido proceso de diversificación de formas que dieron lugar a los primates, entre otras formas. Pero antes de la colisión del asteroide de Chicxulub hace 65 ma los primeros mamíferos coexistieron y compitieron con los dinbosaurios del Jurásico y el Cretácico. En el registro fosil de China se ha descrito la especie Castorocauda lutrasimilis (Qiang et al, 2006), un mamaliforme semiacuático de hace 164 millones de años (Jurásico medio). En el fósil, que está tan bien preservado, se aprecia perfectamente el pelo que cubría al animal y la piel de la cola, que estaba adaptada a la natación y al buceo: era aplanada, ensanchada y escamosa como la de los actuales castores. Los mammaliformes incluyen a los mamíferos y también a otros grupos cercanos, ya extintos, como el de los docodontos, al que pertenece esta especie. Al ser Castorocauda un docodonto muy primitivo, es probable que todo el grupo compartiera esos hábitos acuáticos.
La respuesta biótica a una extinción masiva como la que causó la desaparición de los dinosaurios es la radiación adaptativa. En la transición K/T los grandes beneficiados fueron los mamíferos que experimentaron un rápido proceso de diversificación de formas que dieron lugar a los primates, entre otras formas. Pero antes de la colisión del asteroide de Chicxulub hace 65 ma los primeros mamíferos coexistieron y compitieron con los dinbosaurios del Jurásico y el Cretácico. En el registro fosil de China se ha descrito la especie Castorocauda lutrasimilis (Qiang et al, 2006), un mamaliforme semiacuático de hace 164 millones de años (Jurásico medio). En el fósil, que está tan bien preservado, se aprecia perfectamente el pelo que cubría al animal y la piel de la cola, que estaba adaptada a la natación y al buceo: era aplanada, ensanchada y escamosa como la de los actuales castores. Los mammaliformes incluyen a los mamíferos y también a otros grupos cercanos, ya extintos, como el de los docodontos, al que pertenece esta especie. Al ser Castorocauda un docodonto muy primitivo, es probable que todo el grupo compartiera esos hábitos acuáticos.
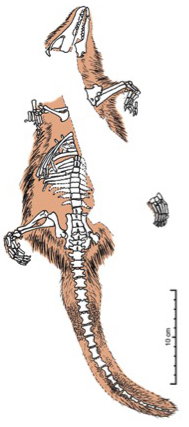
Castocauda lutrasimilis
Similar en proporciones a un ornitorrinco, Castorocauda también alcanzaba su tamaño. Medía casi medio metro, lo cual es un récord en los mamaliformes del Jurásico. Muestra, además, que estos animales no eran todos diminutos insectívoros con aspecto de musarañas, sino que también evolucionaban en otras direcciones y se adaptaban, a pesar de la competencia con los reptiles, a distintos modos de vida. Además, este fosil prueba que el pelo de los mamíferos se originó antes que los propios mamíferos.
Otro mamaliforme significativo anterior a la transición K/T es Repenomamus gigánticus (Hu et al, 2005). Fue un carnívoro depredador del Cretácico que pudo haber consumido pequeños dinosaurios.

Origen y diversificación de los maíferos.
Así, el origen de los Terápsidos, un grupo de reptiles del Paleozoico con algunas características de mamífero, se habrían originado hace más de 250 ma. El origen de la homeotermia se habría dado durante el Mesozoico antes de la divergencia entre los monotremas (mamíferos ovíparos) y los mamíferos placentarios marsupiales y euterios, que a su vez se habría diversificado hace una 150 ma. Por tanto, tanto Castorocauda como Repenomamus són fósiles anteriores la aparición de los mamíferos modernos, e incluso anteriores a la gran radiación de los primates que se dió hace 65 ma. El fosil más antiguo con rasgos claramente atribuibles a un primate es Darwinus masillae, datado en 47 ma. Se encontró en Alemania y es parecido a un lemur actual, pero sus dientes y dedos de los pies difieren, lo que sugiere que se trata de un mamífero primitvo que no presenta las adaptaciones arbóreas características de los primates actuales.
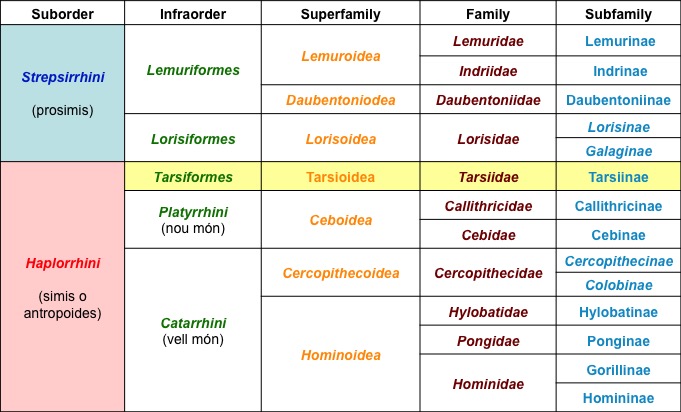
Taxonomñía de los primates actuales.
a. Evolución y diversificación
Los primates son organismos evolutivamente muy generalizados. La reducción del número de piezas dentales es poco marcada en comparación con la mayoría de mamíferos y presentan pentadactilia (5 dedos en manos y pies), un rasgo anatómico poco especializado. En cambio, tienen una clavícula desarrollada que indica especialización a la vida arborícola y la mayoría posee uñas en lugar de garras, que los diferencia de los mamíferos insectívoros arborícolas. Tienen los hemisferios cerebrales bien desarrollados, ojos en posición frontalizada y pulgar oponible. La cola puede no estar presente o no, y puede ser prensil. En general son arborícolas, aunque en grado variable. Presentan glándulas mamarias en posición pectoral y forman grupos familiares de tamaño variable, con un número variable de crías. Presentan actividad sexual continua, exepto lemúridos, con un estro (ovulación) normalmente evidente. Las crías tienen un largo periodo de desarrollo infantil, con una madurez sexual tardía respecto a los demás mamíferos, con una mayor longevidad y conducta social compleja.

Radiación adaptativa de los primates
Sb. O. Estrepsirrhinos: En general de vida nocturna, con ojos grandes. Ausencia de tabique óseo entre fosa temporal y órbita. Anillo orbitario alrededor de la órbita. Ojos separados dirigidos lateralmente. Rápida maduración sexual, con varias crías. Más de un par de glándulas mamarias, útero bicorne. Cara larga con hocico prominente. Presentan rhinario húmedo. Dentición 3/2 I, 1 C, 4/3 Pm y 3 M. Incisivos y caninos inferiores en forma de peine. Labio superior soldado a la encía. La mayoría tiene una uña para limpieza en cada pié.
Infra O. Adapiformes (grupo fósil): Tamaño grande (similares a los lemures actuales). Fórmula dental primitiva (2143). Incisivos inferiores pequeños y verticales. Hocico largo pero ancho. Órbitas con anillo óseo completo.
Infra O. Lemuriformes (Madagascar Islas Comores): Hábitos arborícolas y nocturnos. Comen hojas, frutos e insectos. Patas posteriores más largas que las anteriores. Fórmula dental 2133. varia familias: Lorisidae, Lemuridae, Indridae, Daubentoniidae.
Sb. O. Haplorrhinos (antropoides): Cerebro de mayor tamaño que estrepsirrhinos. Aparato olfativo reducido, ausencia de rhinario. Órbitas frontalizadas y cerradas. Cóndilos y foramen magno adelantados. Dentición 2 I, 1 C, 2/3 Pm, 2/3 M. Capacidad prensil bién desarrollada. Dimorfismo sexual marcado. Craneo grande y redondeado. Labio superior móbil. Rhinario seco y piloso. Ojos en posición frontal. Extremidades anteriores largas.
Infra O. Tarsiiformes: No tienen rhinario. Septo óseo orbitario parcialmente formado. Ojos grandes (se asemeja a los estrepsirrhinos). Tarso largo. Arborícolas, insectívoros y nocturnos. No tienen peine dental. Presentan dos unñas peinadoras en cada pie.
InfraO. Platyrrhini (monos del Nuevo Mundo): Nariz aplanada y ancha. Forámenes nasales separados por una lámina cartilaginos gruesa orientada hacia fuera. La mayoría diurnos. Dos familias principales: Cebidae (arborícolas, cola prensil) y Callitricidae (garras afiladas, cola desarrollada, no tienen M3).
InfraO. Catarrhini (monos del Viejo Mundo): Cinomorfos (hocico largo). Cuadrúpedos terrestres o arborícolas. Callosidades isquiáticas. Dos superfamilias: Cercopithecoidea (Cercopithecidae) y Hominoidea (Hylobatidae, Pongidae y Hominidae).
Primates del Paleoceno (65-57 ma)
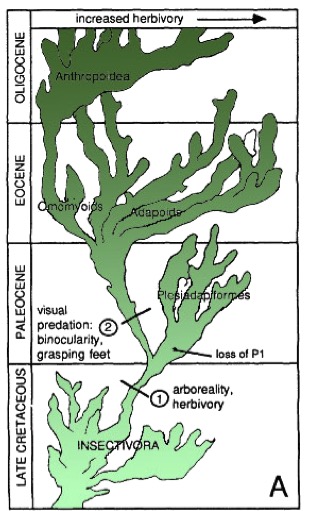
Radiación evolutiva de los primates.
En el Cretáceo final desaparecen los dinosaurios y el Paleoceno es la Era de los Mamíferos. Los primates primitivos evolucionaron a partir de los mamíferos insectívoros. Los primates, musarañas, lemures voladores y murciélagos están estrechamente emparentados (J Hum Evol 16, 1-20, 1987).
Purgatorius (-/3143)
En el Paleoceno inicial el género Purgatorius era un pequeño primate de unos 150 gramos de peso con una fórmula dental primitiva (-/3143) y algunos carateres parecidos a los de primates posteriores: incisivos centrales grandes, último premolar molarizado, trigónido bajo, talónido ancho y último molar alargado. Su alimentación era insectívora (como sugiere su pequeño tamaño y la altura de las cúspides de los molares), pero muchos caracteres dentarios de los molares indican un cambio de mecánica masticatoria de verticalizada (para comer insectos) a lateralizada (que habría permitida una dieta más omnívora). Este fosil Se encuentra en yacimientos de Norteamérica.
Plesiadapis (-/2133)
En el Paleoceno medio y final se da la primera radiación de los primates arcaicos: los Plesiadapiformes. Es un amplio grupo de primates del Paleoceno y Eoceno inicial en Norteamérica y Europa. Llegan a ser muy numerosos y con gran diversidad taxonómica (más de 25 géneros y 75 especies). Se conservan sobre todo dientes y mandíbulas. El género más conocido es Plesiadapis. Presentan molares con cúspides bajas, molares inferiores com el trigónido bajo y talónido aplanado, M3 no reducido y con talónido extendido. Su fórmula dental es 2133/2133 (más reducida que la de Purgatorius -/3143), por lo que no son antecesores de los Prosimios (que tienen 4 Pm). Su gran variabilidad dental sugiere tenían una gran diversidad alimentaria y ecológica (tanto insectívoros como folívoros), con formas tanto arborícolas como terrestres. Probablemente eran nocturnos o crepusculares. La similitud en algunos caracteres entre plesiadapiformes y roedores sugiere una competencia directa entre estos grupos, lo que pudo ser la causa del declive de los plesiadapiformes en el Eoceno. Las particularidades del grupo lo apartan de la linea evolutiva hacia los prosimios del Eoceno.
Primates de Eoceno (57-37 ma)
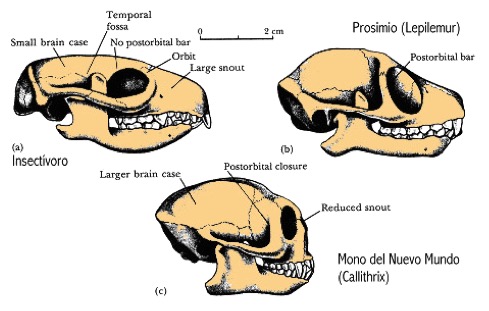
a) mamífero insectívoro sin separación entre la cavidad ocular y la fosa temporal; b) prosimio con anillo orbitario delimitando la cavidad ocular; c) haplorhini con la cavidad ocular cerrada.
En el Eoceno se produce un declive de los Plesiadapiformes. Aparecen las primeras formas parecidas a los actuales Prosimios. El clima es más calido que en el Paleoceno: tropical en Norteamérica y en zonas próximas al círculo Ártico. Los primates de Eoceno tienen todas las características de los primates actuales: hocico más corto que los Plesiadapiformes, barra ósea postorbitaria formando un anillo orbitario (ausente en los mamíferos insectívoros), cerebro más grande y redondeado, zona timpánica similar a los prosimios, dedo gordo del pié oponible y uñas en lugar de garras en casi todos los dedos. Utilizaban más la vista y menos el olfato, y tenían mayor habilidad manipulativa, locomoción más acrobática y saltadora. Tuvieron un éxito desigual en Asia y África respecto a Europa y Norteamérica donde eran abundantes. No se encuentran en Sudamérica ni el la Antártida. Los representantes más antiguos aparecen simultáneamente en Norteamérica y Europa, cuyas faunas del Eoceno inicial son virtualmente idénticas. En Asia son algo posteriores.
La familia más representativa es la de los Adapidae, que en muchos caracteres son los primates más primiticvos, de tamaño relativamente grande (similares a los lemures actuales). Su fórmula dental es primitiva, con cuatro premolares (2143/2143) e incisivos inferiores pequeños que están dispuestos verticalmente (a diferencia de los lemures actuales). Se distinguen diversas subfamilias: Notharctinae (Norteamérica y algo en Europa, cinco géneros, el más antiguo Cantius (2143/2143); Adapinae (Europa, con un género tardío de Norteamérica, 2143/2143, algunos 2133), la especie más conocida es Adapis parisiensis del Eoceno final que desapareció con el cambio climático (a frío) del límite Eoceno-Oligoceno (Grand Coupure); Sivaladapinae, tras la desaparición de los notharctinos y los adapinos, aparecen algunas formas en el Mioceno de Asia (India, Pakistan y China), la especie más conocida es Sivaladapis nagrii de la India (2133/2133). LosAdapiformes son muy parecidos a los Strepsirhinos (especialmente a los lemures pero sin peine dental). Ambos grupos retienen la condición primate ancestral. Los estrepsirhinos actuales han conservado durante 60 millones de años la morfología de los adapiformes.

Evolución de los primates desde el Paleoceno al Mioceno.
La familia Omomyidae aparecen en el Eoceno inicial en Norteamérica, Europa y, posiblemente, Asia. En Europa (con un solo género) presentan menor diversidad que en Norteamérica. Las formas primitivas recuerdan a los primeros adapiformes. Su fórmula dental era 2143/2143 pero enseguida desaparece un premolar (2133/2133), lo que indica que la pérdida de un premolar en los plesiadapiformes anteriores era independiente de la de los omomíyidos, más evolucionados. Sus craneos recuerdan a las társidos y gálagos, ya que son cortos y con grandes órbitas. Tres subfamilias: Anaptomorphinae (primitiva, Teilhardina en Europa y Norteamérica en el Eoceno inicial, los demás son todos de Norteamérica; muchas especies tienen dos premolares 2123/2123; el cráneo sólo se conoce en el género Tetonius); Omomyinae (insectívoros, frugívoros y folívoros, se conoce el cráneo Rooneya y el postcraneal de Hemiacodon gracilis); Microchoerinae (Europa, derivan de formas de anaptomorphinae, Formas saltadoras similares a Tarsius actual; alguna evidencia de omomyidos en Asia, las formas que recuerdan claramente a los társidos se encuentran en el Eoceno en los depósitos de Fayun en Egipto, Afrotarsius chatrathi del Oligoceno inicial, y en Tailandia Tarsius thailandica, del Mioceno). Desde su aparición los primeros prosimios se pueden clasificar en dos grupos: los adapiformes (parecidos a los actuales lemúridos) y los omomyidos (similares a los tarseros y gálagos). Los primeros representantes de ambas familias son muy similares, indicando una divergencia reciente (quizá en el Paleoceno final o Eoceno inicial). Los dos grupos tuvieron una gran radiación adaptativa de las que actualmente quedan representantes estrepsirrhinos en selvas de África, Madagascar y Asia.
Primates de Oligoceno (37-23 ma)
En el Oligoceno la distribución de los continentes era similares a la actual, excepto en la conexión entre Norteamérica y Sudamérica. La India cerraba ya el mar de Tethis y Sudamérica y Australia estaban separadas de la Antártida. Ello creó nuevas corrientes alrededor de la Antártida, causando la caida de temperaturas, glaciación de los polos y bajada del nivel del mar, especialmente en el Oligoceno medio (hace unos 30 millones de años). El registro fósil de primates del Oligoceno cambia drásticamente. Los prosimios del Eoceno desaparecen en Europa y son raros en Norteamérica. Se hacen más abundantes en África y se encuentran por primera vez en Sudamérica (entrada desde África). Las formas más antiguas de primates superiores son dos especies poco conocidas del Eoceno final: Amphipithecus mogaungensis y Poundangia cotteri. Recuerdan más a los primates superiores que a los adpiformes y omomiyidos. Pero es del yacimiento de la depresión de Fayum en Egipto de donde procede la mayoría de fósiles del Oligoceno que conocemos. Los sedimentos de la formación de Jebel Qatrani indican clima templado, seco y algo estacional; la vegetación era similar a la de las selvas tropicales del Sudeste asiático con zonas pantanosas. Se distinguen dos familias:
- F. Parapithecidae: son los más primitivos de todos los primates superiores (antropoideos) conocidos y presentan caracteres que los diferencian del resto de primates del Viejo Mundo. Tienen una fórmula dental primitiva (2133); los incisivos inferiores son pequeños y espatulados. Los molares tienen cúspides bajas y redondeadas. Se han descrito tres géneros: Qatrania es el más primitivo (de pequeño tamaño, comía frutos más que insectos); Apidium (con dos especies) tiene caninos dimórficos, comía frutos y semillas, era un buen saltador; Parapithecus (con dos especies) es enigmático y poco conocido. Su posición filogenética está en la base de los modernos Platirhinos pero sus similitudes son probablemente caracteres antropoides primitivos retenidos tanto en platirhinos como en catarhinos, con los que los parapithecidos comparten caracteres ancestrales.
-

Aegyptopithecus zeuxis
F. Propliopithecidae: antroppoides de gran tamaño (2123/2123), parecidos a los antropoides posteriores, pero son más primitivos que cualquier catarrhino actual. Propliopithecus y Aegyptopithecus zeuxis tienen una morfología craneal primitiva pero más avanzada que la de cualquier prosimio del Eoceno. Comparten algunos caracteres con los Platyrhinos y con los omomyidos de África. El primer registro de platyrhinos en el registro fósil de Sudamérica proceden del Oligoceno final (5 a 10 ma). Los depósitos Paleocenos y Eocenos de Sudamérica son abundantes y contienen mamíferos únicos de este continente, lo que refleja el aislamiento del resto de masas de tierra. Sin embargo, no contienen primates. En Sudamérica sólo aparecen Primates en el Oligoceno final junto con otros mamíferos del Viejo Mundo (roedores). Sudamérica era una isla en el Oligoceno. Pero existieron áreas de aguas poco profundas y rosarios de islas en el Atlántico Sur y en periodos de bajo nivel del mar (Oligoceno medio) las placas continentales de África formaban probablemente tierra seca emergida, reduciéndose las distancias de mar abierto. La reconstrucción de las corrientes marinas parece favorecer el paso desde África. Las formas fósiles de África son sin duda precursoras de los platyrhinos (formas antropoideas de Fayum). Se ha propuesto que llegaron por “rafting” en masas emergidas flotantes con vegetación.
Primates del Mioceno (23-5 ma)
En el Mioceno las temperaturas se vuelven más cálidas. Cambios tectónicos causaron el cierra del mar de Tethis y el surgimiento de la península arábiga. Africa, en el Mioceno inicial, estaba aislada; en el Mioceno medio (18-12 ma) se unió por la Peninsula Arábiga y por Gibraltar con Eurasia. En el Mioceno final, al menos en una ocasión, el mar de Tethis se seca completamente 6 ma y estuvo así hasta que la cuenca se rellenó de nuevo desde el Atlántico y se formó el Mediterráneo actual. La India continúa su choque con el continente asiático, formando el Himalaya y se formaron también los Alpes, los Pirineos, la cordillera del Atlas (Norte de África) y los Montes Zagros (Próximo Oriente, al sur de Irán). En África del Este el Mioceno se caracterizó por una actividad volcánica considerable junto con la formación del Valle del Rift, donde (en los sedimentos del Mioceno inicial de Kenya y Uganda) se encuentran los más antiguos fósiles de primates del Viejo Mundo y gran cantidad de Antropoides que iniciarán la radiación de los Hominoidea.
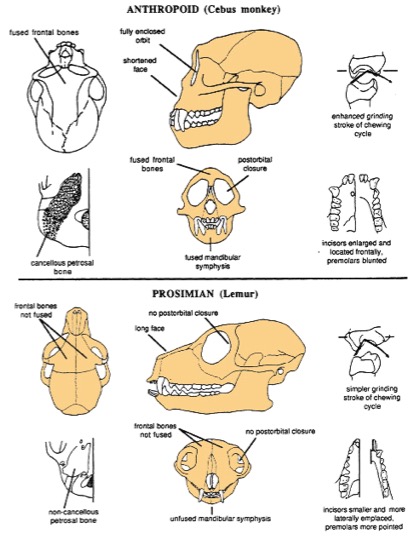
Diferencias anatómicas entre prosimios (abajo) y antropoideos (arriba). Los rasgos especializados de los antropoideos son: cierre de la sutura metópica, cara corta, órbita completamente cerrada, hueso petroso esponjoso, sínfisis mandibular fusionada, capacidad trituradora del ciclo masticatorio, incisivos grandes y premolares redondeados.
A comienzos del Mioceno hay documentadas glaciaciones en la Antártida y en Islandia y a finales del Mioceno, hace 5-6 ma, se produce una clara reducción de los bosques tropicales. En Europa se extinguen los hominoideaos por la presión del frío en este momento. En los sedimentos del Mioceno inicial y medio de Kenya y Uganda hay evidencias de la radiación de los primeros antropoides: los proconsúlidos, más avanzados que Aegyptopithecus y Propliopithecus del Oligoceno inicial (Fayum). Los proconsúlidos tienen todos los caracteres de los catarrhinos actuales y su tamaño varía desde unos 3 Kg de peso (Micropithecus) hasta unos 50 Kg (Afropithecus y Proconsul). Ocuparon un rango ecológico similar al de los monos del Viejo Mundo y hominoideos actuales, desde bosques tropicales a bosques abiertos, con quienes comparten caracteres dentales (2123/2123).
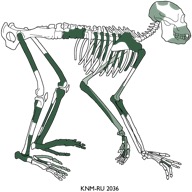
Proconsul africanus
La especie Proconsul africanus es la especie más generalizada de este periodo. Su esqueleto postcraneal sugiere que el aparato locomotor sería bastante primitivo. Carece de los caracteres especializados de la rodilla y el codo de los catarrhinos y por ello su locomoción sería más versátil. En el Mioceno inicial se han descrito varias especies: P. africanus, P. major y P. nyanzae (que difieren básicamente en tamaño), Afropithecus (dificil de asignar); Turkanapithecus, Rangwapithecus (de selva lluviosa); Nyanzapithecus (con caracteres dentales que lo asocian con Oreopithecus), Limnopithecus (arborícola cuadrúpedo), Dendropithecus (el más suspensor de todos, con extremidades largas y finas parecidas al mono araña actual), Simiolus (parecido a Rangwapithecus), Micropithecus clarki (la especie más pequeña) y Dionysopithecus (de China). La especie Equatorius africanus es posterior a los homínidos del Mioceno temprano, pero anterior en el tiempo a la especie Kenyapithecus, por lo que se le considera un miembro tardío de la radiación de los homínidos en el Mioceno medio en África. Kenyapithecus, del Mioceno medio, estaría asociado a hábitats secos y abiertos. Difiere también de los otros por tener el esmalte de los molares más grueso y premolares superiores más grandes. Kenyapithecus parece estar más cerca de la radiación de los antropoides y humanos que los otros géneros, incluido Afropithecus. La diversidad de los primates del Mioceno refleja la diversidad de hábitats que ocuparon: desde selvas tropicales cerradas (Limnopithecus, Micropithecus y Proconsu major) hasta zonas de bosque seco (Dendropithecus, Proconsul africanus y Proconsul nyanzae), con sistemas de locomoción cuadrúpeda, saltadora, suspensora y terrestre. Los Proconsúlidos parecen ser los primeros antropoides diferenciados de los cercopitécidos actuales.
Monos del Viejo Mundo
Los precursores de los monos del Viejo Mundo aparecen a la vez que los antecesores de los antropoides, en el Mioceno inicial. Su radiación tuvo lugar con posterioridad. Desde el Plioceno hasta la actualidad el registro fósil de los catarrhinos del Viejo Mundo es muy abundante, tanto en África, Europa como Asia.
- F. Victoriapithecidae: el primer registro de cercopitécidos procede del Norte y Este de África, en el Mioceno inicial. Son más primitivos que los representantes actuales. Dos géneros descritos: Prohylobates (Mioceno inicial de Egipto, Libia y Norte de Kenia) y Victoriapithecus (Mioceno medio en la isla de Maboko del Lago Victoria en Kenia y un diente del yacimiento del Mioceno inicial de Napak en Uganda) que son muy parecidos. Ambos son eslabones entre los catarrhinos primitivos y los cercopitecoideos modernos.
- F. Cercopithecidae: después de la familia de los victoriapitécidos hay un vacío en el registro fósil de unos 10 millones de años, aunque en el Plioceno y Pleistoceno, el registro fósil es abundante, tanto en África como Asia. La radiación de este grupo es ya muy similar a las especies actuales. Se distinguen dos subfamilias: Cercopithecinae (macacos, banuinos y geladas) y Colobinae (colobinos tanto de Europa, Asia, África).
Hominoidea de Eurasia
Son muy escasos. Cuatro grupos: pliopithécidos, oreopithécidos, dryopithécinos y pónginos
- F. Pliopithecidae: Pliopithecus es el más antiguo y primitivo de los antropoides europeos. Antecesor del gibón pos sus caracteres faciales, pero presenta otros (cráneo y esqueleto) poco especializados. Sería uno de los últimos miembros de la radiación de los catarhinos. El género Laccopithecus, del yacimiento de Lufeng en China, es un antropoide de gran tamaños (12 Kg) virtualmente idéntico a Pliopithecus en la dentición, pero es de mayor tamaño y más robusto en algunos caracteres del cráneo (zigomático). Es el último superviviente de esta familia.
- F. Oreopithecidae: Europeo, 1 sola especie Oreopithecus bambolii en el Norte de Italia. Signos de suspensión arborea: tronco corto y ancho, brazos largos, dedos finos y gran mobilidad de las articulaciones. Oreopithecus se habría originado a partir de los proconsúlidos del Este de África y estaría relacionado con los antropoides actuales.
- F. Pongidae: El género Dryopithecus procede de Europa (y posiblemente de China) y Sivapithecus del Oeste y Sur de Asia. En Grecia Graecopithecus (= Ouranopithecus) difiere en la morfología nasal de Dryopithecus de Trukía y Pakistán. Otros restos de China inicialmente clasificados como Sivapithecus han sido renombrados como Lufengpithecus ya que parecen tener más afinidades con Dryopithecus que con Sivapithecus. Gigantopithecus es un género próximo a Sivapithecus. Era el primate más grande que ha existido. Gigantopithecus blacki perdura hasta el Pleistoceno.
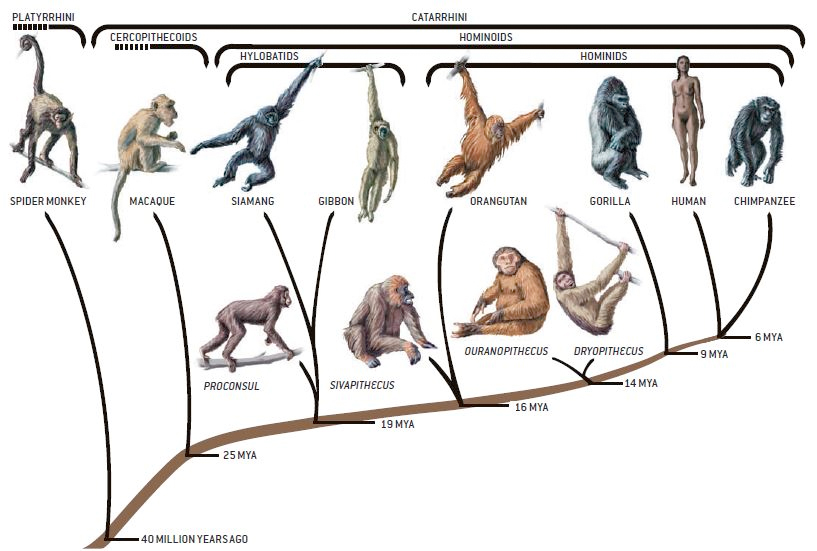
Algunos representantes fósiles de los primates hominoideos actuales.
Estos grupos representan la base del origen de los antropoides actuales (gibón, orangután, gorila, chimpancé y humanos). La diversificación de los gibones se habría producido hace entre 12 y 20 millones de años a partir de catarhinos del Mioceno inicial, peo no está claro su origen. El orangután derivaría de un linaje de Sivapithecus del Mioceno final hace unos 9 a 12 ma (quizá hasta 16 ma). A Proconsul se consideró durante mucho tiempo un buen antecesor pero actualmente se le considera demasiado primitivo y generalizado. La divergencia de los antropoides africanos se habría producido hace entre 6 y 10 ma. El mejor candidato es Kenyapithecus (Samburu Hills) pero se conocen pocos restos. Por lo que respecta a los Homininae, se había considerado a Ramapithecus como nuestro ancestro, hoy se ha asimilado al genero Sivapithecus.
Hominoidea de África
La evidencia más antigua de homininae son fragmentos de mandíbula de 9 ma en Samburu Hills, con rasgos de gorila y capa gruesa de esmalte, que podría ser un carácter ancestral de los homininae. Destacan dos mandíbulas en Kenia: yacimientos de Lothagam de 5.6 ma y Tabarin de 5 ma. Su morfología es muy parecida a la de Australopithecus afarensis pero no se pueden diferenciar de sus antecesores comunes con el chimpancé.

Primates fósiles del Mioceno, en negro las especies asiáticas y en verde las africanas.
- F. Proconsulidae: caracteres catarrinos ancestrales en cráneo y dentición; pero está asociado a los Hominoidea por el postcraneal (no tenía cola).
- F. Hylobatidae: Los representantes mejor conocidos de Hylobatidae son los fósiles de hace 17 ma de Buluk (Afropithecus) en Africa y los de Add Dabtiyah (Arabia Saudí), con Heliopithecus (17-16 crones).
- F. Hominidae: De 17 a 12 ma, representa la segunda radiación de Hominoidea con diferentes rasgos evolutivos
- Sub. F. Afropithecinae: sólo africana, Afropithecus turkanensis (Lago Turkana, Buluk 17 ma), Heliopithecus leakeyi (Península de Arabia, Add Dabtiyah, también de 17 ma), Kenyapithecus africanus (de Africa del Este en Maboko Island, de 15 ma), Otavipithecus namibiensis (13 ma, descubierto en Africa del Sur).
- Sub. F. Kenyapithecinae: Africa del Este y Eurasia. Kenyapithecus wickeri (Fort Tenan, 14 ma) Griphopithecus alpani de Turquía (Pasalar y Candir), así como en Checoslovaquia y en Austria. Todos entre 15-14 crones. Son los restos euroepos más antiguos de hominoidea. Capa gruesa de esmalte. Podría ser este carácter ancestral compartido por ponginae y homininae.
- Sub. F. Dryopithecinae: sólo europeos, entre 12 y 8 ma en Francia, Austria, Alemania, España y Hungría: Dryopithecus fontani (12 ma, Seu d’Urgell y Saint Gaudens), Dryopithecus laietanus (11-9.5 ma, Can Llobateres), Dryopithecus brancoi (11-10 ma, Rudabanya, Hungría, ∼ Rudapithecus), Dryopithecus crusafonti (11-9 ma, Can Ponsic). Todos se extinguieron hace 9 ma por el enfriamiento del clima. Los Dryopithecinae son un grupo heterogéneo del que no se conocen bien sus relaciones filogenéticas.
- Sub. F. Ponginae:
- Sivapithecini: Sivapithecus 12.5-7 ma. El antiguo Ramapithecus se considera hoy la hembra de Sivapithecus. Sivapithecus indicus (Pakistan) y Sivapithecus sivalensis (India), se encuentran desde Turquía hasta Pakistán y la India. Bosque subtropical.
- Gigantopithecini: del Mioceno final al Pleistoceno medio. Gigantopithecus bilaspurensis (India, dientes de 9-6 ma y mandíbulas de 8.5 ma) y Gigantopithecus blackii (Pleistoceno inicial-medio, China).
- Lufengpithecini: Lufengpithecus lufengensis del Mioceno final (China).
- Sub. F. Homininae: capa de esmalte gruesa en molares (Proconsul-Afropithecus-Kenyapithecus), dieta dura por desecación y estacionalidad; cambios en el aparato locomotor (Procónsul-Afropithecus-Dryopithecus-Sivapithecus) hacia la braquiación suspensora. No hay buena evidencias claras para identificar la separación del chimpancé y los primeros hominini. Hay varias hipótesis: Graecopithecus freybergi (10-9.5 ma) que tiene la capa de esmalte más que los australopitecos robustos y aparece en el Este europeo tras la extinción de Dryopithecus en el Oeste; Dryopithecus fontani (12 ma), que tiene el postcraneal muy parecido al del chimpancé, gorila y humanos; Otavipithecus namibiensis, fósil que se sitúa entre Keniapithecinae y Ponginae, se conoce una mandíbula de 13 ma, un frontal de 12 ma similar al del chimpancé (sinus frontal) que documentaría la separación de gorila/chimpancé en 12 crones.
b. Los primates Hominoideos
La Sp. F. Hominoidea (hominoideos) incluye todos los antropoides vivos y extintos, los proconsúlidos (fósiles), hylobátidos y homínidos, que a su vez incluye a los Ponginae. Gorillinae y Homininae (tribu panini y tribu hominini). La Sistemática de los hominoideos se establece en base a la anatomía comparada de las especies vivas, el análisis de especímenes fósiles de un registro escaso, la Sistemática molecular (hibridación, secuencias DNA) y las similitudes evolutivas entre las especies.

Taxonomía de los primates.
Aún así, la taxonomía de los primates hominoideos ha variado considerablemente. Actualmente sabemos que humanos y chimpancés están más próximos entre sí que respecto a cualquier otro primate hominoideo, A pesar de las grandes similitudes anatómicas entre chimpancé y gorila. Dado que la taxonomía de un grupo debe reflejar su filogenia, es lógico que clasifiquemos a los panini y hominini en un mismo grupo (Sb. F. Homininae), que junto a los gorilas (Sb. F. Gorillinae) y los orangutanes (Sb. F. Ponginae) forman la F. Hominidae (Great Apes). Por contra, los gibones (F. Hylobatidae) forman un grupo aparte (Lesses Apes). Así, los humanos pertenecemos a la tribu de los homininos dentro de los homínidos.
Los hominoideos son primates catarhinos (Monos del Viejo Mundo) de tamaño grande, diurnos, que viven mayoritariamente en bosques, con una dieta frugívora y folívora, pero con un cierto aporte de proteinas de origen animal. El modelo reproductor de los hominoideos es poligínico (un macho con un haren de hembras), aunque el número de hembras puede variar considerablemente.
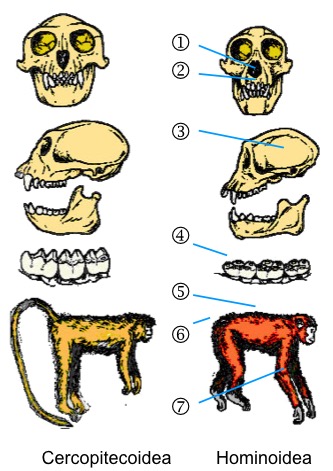
Caracteres derivados de los primates hominoideos. 1. nariz ancha, 2. paladar ancho, 3. volumen craneal grande, 4. dentición bunodonta, 5. columna vertebral flexible, 6. región lumbar corta y ausencia de cola, 7. extremidades anteriores largas.
Los caracteres primitivos compartidos (simplesiomorfías) por todos los representantes del grupo (compartidos por todos los catarhinos) son especialmente evidentes en la dentición. La condición primitiva consiste en tener dientes con cúspides bajas y redondeadas (bunodoncia), auqnue los primates cercopitecoideos muestran una condición derivada (bilofodoncia) que comporta una reducción del número de cúspides y la presencia de crestas. Los hominoideos, en cambio, mantienen la condición ancestral. Pero la mayoría de características anatómicas de los hominoideos son derivadas (apomorfías). Por ejemplo, son especies adaptadas a la braquiación (colgados de las ramas de los árboles), por lo que no tienen cola, tiene los brazos más largos que las piernas y un menor número de vértebras lumbares comparados con los cercopitecoideos. Además, el hueso sacro está expandido, el tórax es ancho con la escápula situada en posición dorsal como es típico de los animales ortógrados (no lateral como en los primates pronógrados) y tienen una cintura pélvida ancha. Además, la capacidad craneal de los hominoideos es significativamente mayor que la de los cercopitecoideos, rasgo que está asociado con una estructura social más compleja. un largo periodo de gestación y lactancia, mayor desarrollo infantil y mayor longevidad.
En caunto a los caracteres dentales, la fórmula dental delos catarhinos es 2123 (2 incisivos, 1 canino, 2 premolares y 3 molares en cada cuadrante). La condición primitiva de los mamíferos es 2143, mientras que en los prosimios y platirhinos es 2133. Por tanto, la fórmula dental 2123 de los hominoideos representa una reducción 2 premolares en cada cuadrante (8 dientes en total) respecto a la condición primitiva, aunque esto representa muy poco comparado con otros grupos de mamíferos, como los osos hormigueros que no tienen ningún diente. La morfología de los molares, es también primitiva en los hominoideos, pero muestra algunas características derivadas. Los molares superiore tien 4 cúspides redondeadas y bajas, y los inferiores 5 con la porción distal (talónido) extendida con 3 de las 5 cúspides, mientras que en el talón los molares superiores sólo tienen 1 cúspide (hupocono).
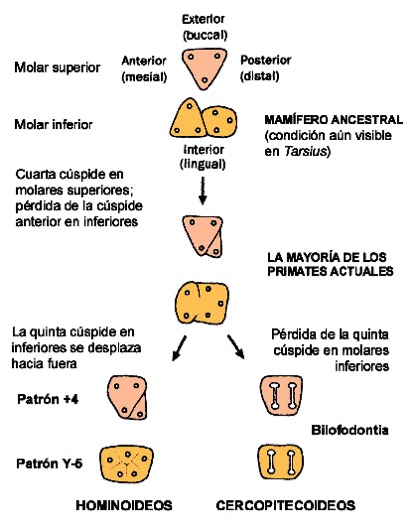
Evolución del patrón morfológico de los molares en primates cercopitecoideos y hominoideos.
Los molares de los primates hominoideos se consideran primitivos ya que conservan las cúspides bajas y redondeadas de los mamíferos ancestrales, pero también tienen alguna condición derivada, como la aparixión del hipocono en el trígono, la reducción de una cúspide en el trigónido y la reordenación de las cúspides en el molar inferior y formar lo que se denomina el patrón Y5. En comparación, los primates cercopitecoideos pierden una cúspide más en el molar inferior y ambos molares presentan una disposición paralela de las 4 cúspides unidad 2 a 2 por crestas en sentido bucolingual (bilofodoncia). Además de la variación en el patrón de cúspides de los molares, los catarhinos comparte una condición derivada en la morfología del canino y el primer premolar respecto a otros primates. Los caninos, especialmente los superiore, son grandes (en particular en los machos) y frecuentemente se observa en la arcada dental un espacio sin dientes (diastema) para que encaje el canino del lado opuesto. Además, el primer premolar inferior (LP3) suele ser sectorial, por tener un sector o cresta de contacto con el borde posterior del canino superior. De hecho, el canino superior contacta tanto con el canino inferior como con el premolar inferior, formando un complejo cortador que está especialmente desarrollado en los primates hominoideos. Sin embargo, los humanos actuales no presentamos estas características ya que nuestro canino es pequeño y no sobresale del resto de dientes en la arcada dental ni tenemos diastema.
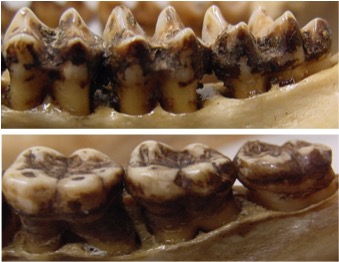
Patrón bilofodonto (arriba) de los molares inferiores de los cercopitecoideos y bunodonto (abajo) de los molares inferiores de los hominoideos.
Las diferencias anatómicas de los molares de los cercopitecoideos (bilofodoncia) y los hominoideos (bunodoncia) reflejan adaptaciones alimentarias distintas. Los cercopitecoideos se han especializado en una dieta eminentemente folívora y las crestas de sus molares sirven para cortar el alimento en trozon pequeños antes de ser ingerido. En cambio, los hominoideos tienen dietas mucho más variadas (omnívoros), desde frugívoras hasta granívoras, y sus cúspides bajas y redondeadas sirven para triturar alimentos duros y abrasivos.
Los humanos modernos compartimos características anatómicas con el resto de primates hominoideos, aunque tenemos muchas especializaciones exclusivas (autapomorfías), en particular las relacionadas con el bipedismo, aunque mantenemos cierta capacidad braquiadora que delata nuestra pertenencia al grupo de los primates hominoideos.
F. Hylobatidae

Hylobates
La famillia de los gibones incluye el género Hylobates con al menos 4 especies (H. sindactilus -antes atribuido al género Synphalangus-, H. concolor, H. hoolock, H. lar). Todos viven en Asia el sudeste asiático (desde la India hasta China, Borneo, Java y Sumatra), ocupando selvas húmedas donde se desplanza por braquición especializada; comen frutos carnosos, hojas tiernas e insectos. Ocupan un territorio relativamente pequeño donde los alimentos están dispersos. Su peso variá entre 5 y 11 Kg y no tienen dimorfismo sexual (ligero en el tamaño del canino). Los incisivos son cortos y anchos y los caninos son largos en ambos sexos, El P3 inferior es sectorial, mientras que el P4 es bicúspide. Los molares son bajos, con una base ancha y cúspides redondeadas. La cara es corta (prognatismo facial reducido) y los machos no tienen crestas en el neurocraneo (sin dimorfismo sexual). La rama mandibular es ancha, como en la mayoría de hominoideos. Los brazos son muy largos (braquiación especializada) y tienen un índice intermembral de 126-147%.Forman parejas estables (monógamos) y los dos progenitores defienden el territorio y cuidan a las crías. Tienen una cría cada 4 o 5 años. El macho del siamang (H. sindactilus) cuida a la cría el segundo año para que la hembra pueda amamantar a otra cría. Las crías permanecen con los progenitores durante unos 10 años.
F. Hominidae
Sb. F. Ponginae

Pongo
La subfamilia ponginae incluye sólo a los orangutanes, con una sola especie (Pongo pygmaeus) aunque varias subespecies separadas geográficamente. Viven el el sudeste asiático (en las islas de Borneo y Java). Ocupan selvas tropicales cerradas y aunque son braquiadores suspensores, su elevado peso les obliga a una locomoción mayoritariamente terrestre, especialmente los machos. Los brazos son largos, con un índice intermembral de 139% y las falanges de las manos curvadas, y las piernas son cortas con el pulgar corto. Son básicamente frugívoros, pero también comen hojas, semillas y corteza. Tienen un marcado dimorfismo sexual, los machos pesar en promedio unos 80 Kg, mientras que las hembras pesan unos 35 Kg. Tiene un prognatismos alveolar muy marcado y el craneo es robusto y con cresta en los machos. La rama mandibular es ancha. El incisivo central superior (UI1) forma una pala muy ancha (característica de una dieta frugívora) y el laterl (UI2( es pequeño. Los caninos son grandes y dimórficos (sobresalen del resto de diente), y tienen un marcado distema en el maxilar superior. Los molares son robustos y presentan una capa de esmalte gruesa que indica que la dieta es abrasiva. Los orangutanes tienen un patrón reproductor poligínico. Los machos son solitarios y controlan un territorio amplio donde viven aisladas una o varias hembras con sus crías (los machos no cuidan a las crías). Los machos se encargan de la defensa del territorio y se desplazan entre los grupos de hembras y crías. La hembra madura sexualmente hacia los 7 años y el macho experimenta una maduración social que varía entre los 8 y los 15 años o más en función de la densidad de grupos y de machos adultos que hay en el territorio
Sb. F. Gorillinae

Gorilla
La subfamilia gorillinae incluye dos especies (Gorilla gorilla, Gorilla beringei) con diversas subespecies en función de la distribución geográfica. Viven en selvas tropicales, tanto en zonas de montaña como en tierras bajas. La dieta puede variar, desde folívoros a frugívoros incluyendo tallos y médula o semillas. Los gorilas presentan un marcado dimorfismo sexual. Las hembras pesan entre 70 y 100 Kg, mientras que los machos varían entre 170 y 299 Kg. La cara es muy prognata y el cráneo robusto, con crestas prominentes en los machos. Los incisivos son pequeños (no es un frugívoro especializado), los caninos muy grandes y con marcado dimorfismo sexual, el primer premolar es sectorial y los molares son bajos y tienen el esmalte fino (a diferencia del orangután). La rama mandibular es muy ancha y robusta. Los brazos son algo más largos que las piernas, que tienen un pulgar largo, y tienen un índice intermembral de 116%, lo que indica que no son braquiadores especializado. Su elevado peso corporal les obliga a desplazarse por el suelo de forma cuadrúpeda. Tienen el pié plano (a diferencia del orangután que lo apoya lateralmente cuando camina por el suelo) y apoyan el dorso de la segunda falange de los dedos de la mano durante la marcha: caminan con los nudillos (knucle-walking), igual que el chimpancé. Los gorilas son poligínicos estrictos, con un solo macho alfa y diversas hembras que forman un harén reproductor. Los machos controlan un territorio de tamaño variable, aunque el grupo se desplaza en busca de alimento. Los machos compiten entre si por las hembras y el territorio. Las crías y los machos inmaduros permanecen en el grupo familiar hasta que maduran sexualmente. Los grupos intercambian hembras (exogamia) y los machos sin harén viven al magen del grupo del macho dominante.
Sb. F. Homininae
Tribuu Panini
La tribu panini incluye dos especies: el chimpancé común (Pan troglodytes) y el bonobo o chimpancé pigmeo (Pan paniscus).

Pan troglodytes
El chimpancé común (Pan troglodytes) comprende varias subespecies (P. t. troglodytes, P. t. verus, P.t. schweinfurthii) que difieren en su distribución geográfica. su hábitat varía desde selvas tropicales hasta sabana árida. Su dieta depende del hábitat ocupado pero los chimpancés son mayoritariamente frugívoros e incluyen otros alimentos dependiendo de la estación del año y la disponibilidad. También consumen pequeños animales que cazan de forma cooperativa y capturan insectos, especialmente termitas, utilizando herramientas. Presentan un dimorfismo sexual moderado, menor que el del gorila. Las hembras pesan unos 30 a 40 Kg y los machos entre 40 y 60 Kg. La cara es corta y el neurocraneo no presenta crestas como el macho del gorila. Los incisivos son amplios y el canino moderado, con dimorfismo sexual. Los premolares son cortos, con el talón poco marcado, y los molares son redondeados y con el esmalte fino (igual que el gorila). El índice intermembral es de 103 a 106%, menor que el del gorila, lo que indica una locomoción terrestre más que braquiador. Las piernas son cortas y se desplaza por braquiación en los árboles y con los nudillos (knucle-walking) en el suelo como el gorila. Su estructura social es bastante compleja. Forma grupos familiares poligínicos (un macho dominante con varios machos emparentados, diversas hembras y las crías) de menor tamaño que el gorila. Estos grupos se puede unir (fusión) o separar (fisión) en función de la disponibilidad de recursos t la estacionalidad anual (sistema de fusión-fisión con exogamia de hembras). Los machos del grupo defienden el territorio, que es amplio, y las hembras interactúan más com otras hembras y las crías que con los machos. Se trata de un patrón reproductor más laxo que el del gorila, con varios machos emparentados que cooperan en la defensa del terrotorio y del grupo familiar.

Pan paniscus
El bonobo (Pan paniscus) difiere considerablemente del chimpancé común. Vive en selvas tropicales de África Central al sur del río Zaire. Su dieta es muy variada pero se basa mayoritariamente en el consumo de frutos maduros y también hojas, insectos y pequeños animales que caza. Presentan un dimorfismo sexual muy reducido. Las hembras pesan en torno a los 30 Kg y los machos unos 45 Kg. Los incisivos son anchos, los caninos relativamente pequeños con escaso dimorfismo sexual, el primer premolar es ligeramente sectorial y los molares reducidos y con esmalte fino. La cara es corta (prognatismo reducido), el neurocraneo no presenta crestas y la mandíbula es bastante gracil. Los brazos son relativamente cortos, con un índice intermembral de 102%. Se desplaza por braquiación en los árboles y es cuadrúpedo en el suelo, dondes se desplaza apoyando los nudillos (knucle-walking). También practican con cierta frecuencia un bipedismo inestable para realizar diversas actividades (en mayor grado que el chimpancé común). El patrón social es de fusión-fisión en función de los recursos, pero la sociedad del bonobo es matrilineal (a diferencia del chimpancé común que es patrilineal). Los machos de un grupo no necesariamente están emparentados y cooperan en la búsqueda del alimento. Además, todos los machos se pueden reproducir dentro del grupo y se utilizan las relaciones sexuales para evitar la competencia y reducir tensiones. Las hembras cooperan intensamente entre si y establecen relaciones de amista para la búsqueda de alimento y el cuidado de las crías.
Tribu Hominini

Homo sapiens
La tribu hominini actualmente incluye sólo la especie Homo saliens. Originalmente ocupó bosques de galería y sabanas abiertas en África, pero ahora ocupa todos los hábitat posibles con ayuda de su compleja tecnología. Presenta un marcado dimorfismo sexual. Las hembras pesan unos 60 Kg en promedio y los machos unos 85 Kg. La cara es muy corta y no tiene prognatismo facial. El craneo es muy gracil y la rama mandibular es reducida y poco robusta. El incisivo central superior es más ancho que el lateral, los caninos son muy pequeños y no sobresales del resto de dientes (no hay diastema), los dos premolares son bicúspides y cortos, y los molares son pequeños y gráciles, con el esmalte grueso. La dieta es muy variada e incluye frutas, vegetales, insectos y carne. El índice intermembral es menos de 100%, lo que indica que las piernas son más largas que los brazos, lo que indica un bipedosmo especializado, sin dedo oponible en el pie y pulgar largo con una elevada capacidad manipuladora en la mano.Los grupos sociales son complejos. El marcado dimorfismo sexual en tamaño sugiere competencia entre machos y poliginia, con una marcada jerarquía social. El modelo reproductor es en gran medida monógamo, aunque la poligamia está aceptada en algunas sociedades. La conducta sexual humana es compleja y se han hecho algunas comparaciones con la conducta sexual del bonobo.
c. Conducta social
La condición primitiva de la mayoría de mamíferos, y tanbién de los primates, es la poliginia de uno macho alfa dominante que se reproduce con varias hembras. En este caso, se da una fuerte competencia entre los machos para mantener un ‘harén‘ de hemras. Este sistema reproductor asegura que los caracteres más adaptativos son los que se transmiten de generación en generación. La competencia entre machos se puede establecer en base al territorio (el macho controla el territorio donde están las hembras) o mediante un control directo del grupo de hembras. En los grupos poligínicos, ya sea de uno (caso del gorila) o varios machos (como en el chimpancé), las hembras no están emparentadas y el grado de cooperación entre ellas para obtener recursos o cuidar a las crías es variable. La poliandria es una condición derivada poco frecuente. En primates se da en algunos prosimios y en los titís (callitrícidos) sudamericanos. La ventaja de la poliandria (una hembra se reproduce con varios machos) reside en la necesidad de incrementar la tasa de reproducción de la hembra compartiendo las labores de cuidados de las crías con los machos. La monogamia es poco frecuente. Generalmente se interpreta como un caso restringido de poliginia con una sola hembra y suele estar relacionada con escasez de recursos, cuidados parentales largos y compartidos, y reducción de la competencia entre machos. En los hominoideos la monogamia se da en los gobones, que forman parejas estable en un territorio propio.
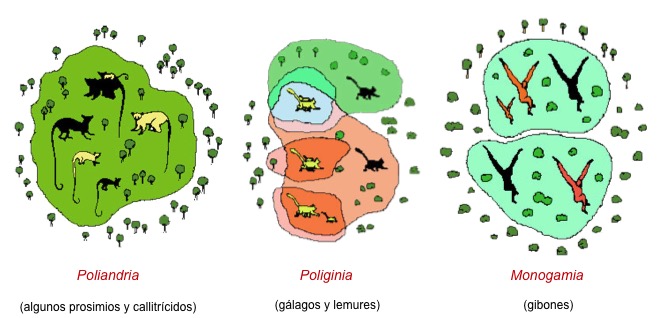
Modelos de estructura social en Primates.
En los primates superiores (cercopitecoideos y hominoideos) la poliginia es el modelo mayoritario, aunque pueden formar grupos de hembras con un solo macho (gorilas y babuinos) o con varios machos emparentados (chimpancés y mandriles). El patrón social y reproductor del chimpancé es especial ya que los grupos pueden dividirse (fisión) o agruparse (fusión) en función de la disponibilidad de recursos. En este modelo los machos compiten por las hembras y las hembras lo hacen por los recursos alimentarios. Si el alimento es pobre energeticamente (hojas, tallos, raices) pero es abundante en el medio, las hebras no sueles establecer relaciones intensas entre ellas y el grupo está disperso. En cambio, si los recursos son ricos energeticamente (frutos maduros) las hembras forman alianzas estables de cooperación.
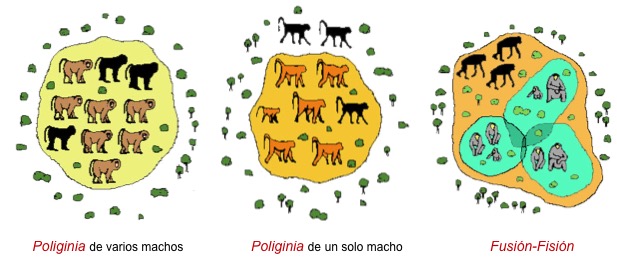
Variabilidad del modelo de poliginia en primates superiores.
El tamaño del grupo de hembras depende entonces de si un solo macho puede defender a las hembras (poliginia de un solo macho), son necesarios varios machos para controlar un grupo grande de hembras (poliginia de varios machos emparentados) o son necesarios los cuidados de ambos progenitores para alimentar y proteger a la cría (monogamia). En los primates hominoideos actuales se da una gran variabilidad de modelos sociales y reproductores, que difieren sustancialmente.
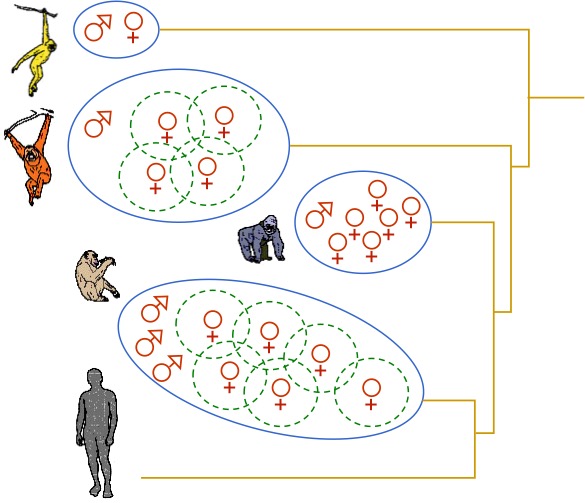
Modelos de estructura social de los primates hominoideos.
Los gibones son monógamos, probablemente como respuesta a la necesidad de proteger y alimentar a la cría. Ambos progenitores se encargan de su cuidado y defienden el territorio en el que compiten con otras parejas. Puede considerarse, por tanto, como un modelo de poliginia de un sólo macho. La ausencia de dimorfismo sexual en los gibones sugiere que la monogamia reduce la necesidad de que los machos compitan por las hembras, al menos de forma directa. Los orangutanes son poligínicos pero la competencia entre machos se establece a través del territorio. Las hembras viven separadas y el macho diminante controla territorios amplios con varias hembras. El elevado dimorfismo sexual del orangután indicaría una gran competencia por la reproducción entre machos de distintos territorios o con machos jóvenes periféricos. La maduración sexual de los machos depende de factores sociales, como la densidad de machos adultos en el territorio o la densidad de grupos y número de hembras. Así, pueden retrasar su maduración sexual hasta que las condiciones de reproducción sean más favorables, permaneciendo más tiempo en el nucleo familiar. Los gorilas practican la poliginia de un solo macho dominante. El gran dimorfismo sexual del gorilla pone de relieve este modelo extremo de estructura social. La competencia entre machos es muy intensa ya que sólo se reproducen los machos más capaces. Como los recursos alimentarios del bosque tropical donde viven son abundantes, no es necesario controlar el territorio y un solo macho es suficiente para controlar un grupo, no muy numeroso, de hembras. El chimpancé común tiene también un patrón reproductor poligínico, pero con varios machos emparentados que cooperan en la defensa del territorio de alimentación. Además, el sistema de fusión-fisión hace que los grupos familiares se unan en temporada de abundancia de alimentos, momento que se aprovecha para el intercambio de hembras entre grupos (exogamia de hembras para evitar la consanguinidad). En cambio, el bonobo tiene un modelo social particular. Forman sociedades matriliniales en las que las relaciones sociales se establecen mediante cooperación y prácticas sexuales cuya función es relajar las tensiones y la competencia. Son grupos muy promiscuos y todos los machos se reproducen. No hay un macho alfa con exclusividad reproductora. De hecho la jerarquía se establece a través de las hembras y los machos la adquieren en función de las relaciones familiares que tiene con ellas.
El dimorfismo sexual está, por tanto, relacionado con la competencia entre machos. Cuanto mayor la competencia entre machos por la reproducciñon con las hembras, mayor dimorfismo. Es el caso del orangután y el gorilla, en los que los caracteres sexuales (tamaño del pene y de los testículos) son relativamente pequeños respeto a su gran tamaño corporal (el pene del gorila es extremadamente pequeño comparado con los demás hominoideos). En cambio, el emnor dimorfismos sexual de los giboes sugiere ausencia de competencia entre machos, quizá causado por su patrón reproductor monógamo. Pero en el chimpancé y humanos el dimorfismo sexual no parece estar directamente asociado al patrón reproductor. Los machos de chimpancés común compiten por las hembras, pero son varios machos los que coopera, y en el bonobo la competencia se ha reducido a través de una conducta sexual compleja (su dimorfismo sexual es menos marcado que el del chimpancé). Sin embargo, la competencia por la reproducción en los machos de chimpancé se ha establecido a través de los órganos sexuales. Los testículos del macho son grandes para poder inseminar a la hembra con una mayor cantifdad de espermatozoides, incrementando así la probabilidad de fecundación en una sociedad más o menos promiscua con varios machos reproductores. Por contra, las hembras presentan los genitales externos extremedamente voluminosos en la época de ovulación (estro) con el fin de limitar la competencia a aquellos periodos de ciclo ovulatorio en que son fértiles.
En humanos el dimorfismo sexual es considerable y no parece haberse reducido por la monogamis (si este es en realidad nuestro patrón reproductor). En cambio, la competencia por la reproduccion (o la selección de pareja) parece haberse establecido en base a los atributos sexuales tanto primarios (genitales) como secundarios (no directamente relacionados con la reproducción, como masa muscular, tamaño del pene, senos o glúteos). Estos caracteres sexuales secundarios sirven de atractivo y ejercen una gran influencia en la selección de pareja reproductora. Sin duda los patrones reproductore de las sociedades humanas son complejos y difíciles de clasificar en categorías simples como monógamo y polígamos. A pesar de ello, se ha intentado encotrar un modelo de conducta social que se pueda aplicar a nuestros ancestros más remotos con el fin de determinar cómo habría sido nuestro patrón social y reproductor primitivo para poded deducir cómo ha evolucionado.
Modelos de especie única
Modelo del babuino de sabana

Babuino de sabana abierta.
El babuino es un cercopitecoideo, por lo que está más lejanamente relacionado genéticamente con nosotros. Aún así, los babuinos constituyen un modelo apropiado para los homininos ancestrales porque comparten un hábitat similar: la sabana abierta. El babuino de sabana vive en grupos de hasta un máximo de 200 individuos. Una banda está formada por hembras maduras (emparentadas), su descendencia y machos maduros no emparentados. Presentan un marcado dimorfismo sexual y establecengrupos poligínicos de un solo macho con múltiples hembras. Este modelo se ha demostrado que es desproporcionado para los primeros homininos, en parte porque en la sabana abierta la competencia con los carnívoros seria muy grande (los carnívoros sociales como el licaones, hienas, leones o guepardos tiene modelos sociales distintos especializados todos ellos para la obtención de proteinas de origen animal), pero también porque requiere la formación de grupos grandes y territorios extensos. Además, actualmente saben¡mos que los primeros homininos evolucionaron el el bosque cerrado y no en la sabana.
Modelo del chimpancé común
Es nuestro más próximo pariente. El comportamiento de las especies es único y las especies vivas no representan el de especies fósiles, por lo que las extrapolaciones al pasado a partir de modelos actuales puede ser engañosa. La dentición robusta de los parantropinos, no conforman el patrón observado en el chimpancé común, que tendría una dieta blanda. A pesar de ello, se ha propuesto el modelo de poliginia de varios machos como modelo social de los homininos en la transición entre el bosque cerrado y la sabana. Pero el patrón de fusión-fisión parece muy especializado para ser aplicado a nuestros ancestros.
Modelo del chimpancé pigmeo
El bonobo (Pan paniscus) es similar en proporciones corporales a los primeros homininos (como A. afarensis). Su sociedad se centra en la hembra y es igualitaria (el sexo es el sustituto de la agresión). La actividad sexual del bonobo es grande y bastante promiscua, copulando frecuentemente de cara, y usualmente como un modo de reducir la tensión del grupo. No es seguro que sea un mejor modelo que el chimpancé común, a pesar de la separación parcial de sexo y reproducción, como ocurre en nuestra especie. Además, el hecho que chimpancé y bonobo difieran en sus modelos de conducta sexual sugiere que el patrón reproductor del bonobo es derivado y que cualquier conducta parecida a la de los humanos habría aparecido independientemente en ambas especies.
Modelos filogenéticos
Se trata de modelos muy conservativos que buscan características básicas del comportamiento compartidas entre los primates hominoideos. Si un comportamiento se encuentra en las cuatro especies de homínidos es probable que también estuviese en el ancestro, dado que de no ser así tendría que haber evolucionado independientemente en cada especie (homoplasia). Por contra, si las cuatro especies difieren respecto a un carácter no se puede inferir el comportamiento del ancestro. El análisis de 14 rasgos del comportamiento de los primates hominoideos (estructura social, interacciones macho-hembra, agresiónes entre grupos, etc.) mostró que 8 eran comunes a gorila, chimpancés y humanos, y 6 no compartidos. De llo se pudo deducir que el ancestro común habría tenido estrechas redes sociales, relaciones sociales de macho dominante hostil con muchas hembras, exogamia de hembras y ausencia de lazos entre ellas. Este modelo choca frontalmente con un hipotético comportamiento monógamo no hostil propuesto para A. afarensis.
Modelos de comportamiento ecológico
Los modelos ecológicos intentan determinar el rango de estructuras sociales que habrían tenido a su disposición los homininos ancestrales, para después determinar cómo dichas estructuras podrían ser alteradas ante ambientes cambiantes. La evolución de una estructura de machos con hembras no emparentadas a hembras emparentadas es muy improbable ya que los pasos intermedios dependen en gran medida de factores ecológicos como la disponibilidad de recursos. Si sabes en qué ambiente vivió el ancestro puedes predecir el cambio social a través de los caminos posibles. El modelo ecológico sugiere que los patrones sociales de los hominoideos vivos actuales constituyen estructuras sociales limitadas que solo tienenen común que ninguno se basa en núcleos con hembras emparentadas (que sí se observa en los cercopitecoideos).
En base a estos modelos, la estructura social más probable del ancestro común de todos los hominoideos sería similar al gorila (macho dominante y machos jóvenes, con hembras y crías). En cambio, la reducción de la selva tropical hacia una sabana más abierta favorece la estructura del chimpancé (machos emparentados, con hembras y crías no emparentadas). En este ambiente más abierto, la presión predatora y la distribución limitada de los recursos favorecería el aumento del tamaño de los grupos, que se habría dado en base a las alianzas de parentesco entre machos. La organización social más probable de los australopitecinos iniciales habría consistido en grupos sexualmente mezclados, con machos emparentados, hembras forzadas a proveerse de recursos en áreas grandes, para encontrar recursos estacionales y limitados, y con lazos y asociaciones estables con machos aislados o grupos de machos. Este modelo está de acuerdo con las condiciones ecológicas existentes pero está en desacuerdo con la interpretación del dimorfismo sexual que se atribuye a Australopithecus afarensis (tan marcado como en el orangután). El dimorfismo equivale a competencia entre machos por las hembras. El 74% sociedades humanas son poligínicas y no monógamas, por lo que el dimorfismo sexual de los humanos modernos indicaría competencia entre machos en nuestro pasado. En humanos los testículos son pequeños, lo que descartaría la competencia en grupos promiscuos de varios machos. También se descarta la monogamia por limitaciones ecológicas de la savana abierta, por lo que sólo quedaría la poliginia de un solo macho en un grupo reducido de hembras.
En cualquier caso, la evolución morfológica de los homininos es compleja y no está necesariamente relacionada con la evolución de la conducta sexual.
Referencias
Hu Y et al (2005) Large Mesozoic mammals fed on young dinosaurs. Nature 433, 149-152.
Qiang J et al (2006) A Swimming mammaliaform from the middle Jurassic and ecomorphological diversification of early mammals. Science 311, 1123-1127.
