La evolución morfológica de los hominini desde la divergencia con el chimpancé se centre en tres aspectos principales: el bipedismo, la encefalización y la reducción del aparato masticador. Sin embargo, las adaptaciones relacionadas con estas características no aparecen a la vez en nuestro linaje, por lo que caracterizar las adaptaciones de los homininos más antiguos no siempre es facil.

Músculos masticatorios
Bipedismo: no está claro que sea la adaptación propiamente humana más antigua. Es cierto que el bipedismo humano es único y no se da en los demás primates, pero en los primeros homininos el bipedismo era bastante primitivo y poco especializado, por lo que se suele hablar de bipedismo facultativo. El bipedismo del género Homo (bipedismo obligado) implica cambios en diversas estructuras anatómicas: el foramen magnum está más avanzado cuanto más bípedo (atrasado en los otros hominoideos); la pelvis se modifica afectando al canal del parto, en humanos el feto snace hacia atrás y la cabeza debe girar (esto no es muy adaptativo y es causa de una mayor mortalidad); las alas iliacas giran lateralmente (están dorsalizadas en los demás hominoideos) para que los músculos laterales permitan mantener el equilibrio; el dedo gordo del pie, que es prensil en loshominoideos no humanos deja de serlo en los bipedos; y el ángulo de inclinación del fémur (ángulo valgo) es más grande y la longitud del cuello cambia.
Encefalización: la corteza cerebral está muy desarrollada en los primates superiores y la proporción de los lóbulos cerebrales varía. En los himanos existen diferencias significativas en las areas de asociación cerebral y los hemisferios cerebrales, conectados por el corpus callosum que coordinan funciones complejas del cerebro. En particular en el cerebro humano se da una fuerte lateralización. En el 95-98% de las pesonas el hemisferio izquierdo es el dominante (en el 5% es el derecho). Además, las áreas de asociación parietal y temporal del cerebro están muy desarrolladas en humanos y mínimamente en el chimpancé y gorila, en particular las áreas de Wernike (área de asociación del lenguaje sintáctico que conecta cortex visual y área motora) y de Broca (respuesta motora del lenguaje). El tamaño del cerebro también cambia a lo largo de la evolución de nuestro linaje, pasando de unos 350 a 1400 cc.
Aparato masticatorio: la dentición y las estructuras asociadas a la masticación sereducen en relación con la alimentación. Estas remodelaciones afectan a la musculatura temporal (apófisis coronoide de la mandíbula hasta la linia crotáfite o la cresta sagital) y al músculo masetero (rama mandibular hasta el reborde inferior del malar y principio del arco zigomático). Su desarrollo depende de la potencia masticatoria observable en las estructuras óseas asociadas. Entre los caracteres dentarios que se modifican están el tamaño de los caninos, la desaparición del diastema y la reducción de los incisivos y los molares.
a. Origen y evolución de los hominini
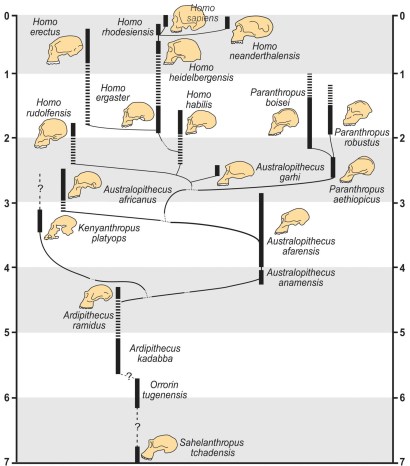
Filogenia de los hominini basada en los caracteres anatómicos de los fósiles.
El análisis de la evolución de nuestro linaje debe respondes a preguntas sobre en qué momento de nuestra evolución aparecen el bipedismo, la encefalización y la reducción del aparato masticador. No siempre será fácil determinar cuál fue el proceso evolutivo preciso y en qué sentido se produjo, pero será necesario determinar el estado (si es primitivo o derivado) de los caracteres que se consideren al realizar comparaciones entre taxones. Las plesiomorfías compartidas (simplesiomorfías) sólo permitirán determinar que los taxones comparados las han heredado de su ancestro cumún, por lo que servirán para agrupar taxones que comparten rasgos primitivos. Las apomorfías exclusivas de un taxón (autapomorfías) nos estarán presentes en otros taxones y, por tanto, no servirán para establecer relaciones de proximidad filogenética. En cambio, las apomorfías compartidas (sinapomorfías) permiten establecer relaciones de ancestro descendiente entre taxones temporales: cronoespecies. Analizaremos nuestra evolución de forma cronológica intentando determinar qué nos define como linaje (los hominini) y nos diferencia del chimpancé (panini), es decir cuáles son las autapomorfías de los homininos. Además, analizaremos las sinapomorfías compartidas por los fósiles para construir la filogenia de nuestro linaje. Existen muchas hipótesis sobre las relaciones ancestro-descendiente en función de los investigadores y sus opiniones y métodos de anñalisis. La filogenia que se muestra en la figura es probablemente la más sencilla y nos basaremos en ella para revisar los taxones más significativos y las relaciones evolutivas entre ellos.
Sahelanthropus tchadensis
Toros-Menalla, Chad (7-6 ma)

Sahelanthropus tchadensis
En el yacimiento de Toros-Menalla en en Chad (datado en el Mioceno final, entre 7 y 6 ma) se encontraron diversos fósiles, incluyendo un craneo (Toumaï, TM 266-01-60-1), un fragmenteo de mandíbula con algunos dientes (TM 266-02-154-1), un canino inferior (TM 266-02-154-2), un incisivo (TM 266-01-448) y la corona de un M3 izquierdo (TM 266-02-154-3) (Brunet et al. 2002, Nature 418, 145–151). El craneo se asignó a la especie Sahelanthropus tchadensis y existe un gran debate sobre su atribución al linaje de los homininos. La reconstrucción virtual del craneo parece confirmar la presencia de muchos rasgos compartidos con los hominini, incluyendo algunos que sugieren que era un hominoideo bípedo. La morfología de la dentición muestra homologías morfológicas en las raícesen los homininos más primitivos, carazcterizada por una reducción de su tamaño y número en los premolares (Emonet et al., 2014, Am J Phys Anthropol 153, 116–123). Además de en la localidad TM 266, también se han descubirto restos de este taxón to en localities TM 247 y TM 292. El especimen TM 292-02-01 es un fragmento mandibular, TM 247-01-02 is un fragmento de cuerpo mandibular y TM 266-01-462 es un fragmento de un P3 superior.
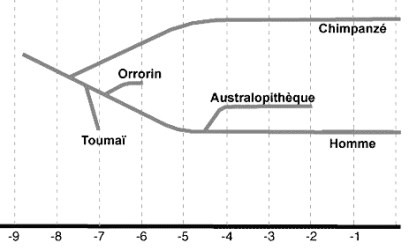
Posición filogenética de Sahelanthropus tchadensis
La comparación del craneo de Sahelanthropus tchadensis con el de los hominoideos africanos sugiere que Toumaï comparte caracteres primitivos con los chimpancés pero en general es más parecido a Australopithecus, en particular en la base del craneo. Sin embargo, Toumaï tiene una cara superior vertical con la región subnasal muy corta. Tiene un volumen craneal de 360-370 cc (parecido al del chimpancé), pero tiene algunas sinapomorfías de los homininos, como un plano nucal, más aplanado, largo y horizontal que el chimpancé, que lo tiene más curvado, una base del occipital corta y un foramen magnum más avanzado. Su dentición es similar a la de Ardipithecus y otros homininos posteriores y tiene un complejo C/P3 reducido con esmalte grueso en los dientes posteriores. Si embargo, algunos aspectos faciales difieren considerablemente de los que presenta cualquier Australopithecus conocido, como la corta región subnasal, las proporciones del paladar, la cara superior verticalizada o la gran robustez del torus supraorbitario, que es continuo, y pueden considerarse caracteres derivados (autapomorfías). En conjunto, Toumaï es un mosaico de caracteres primitivos y derivados. El análisis fenético (comparación de caracteres anatómicos) agrupa a Sahelanthropus con los homininos posteriores, incluyendo Australopithecus, Paranthropus, and Homo, antes que con Pan, principalmente por sus similitudes en la base del craneo y el neurocraneo con los homininos. A pesar de ello, los caracteres derivados de Toumaï sugieren que no es el ancestro directo de los homininos posteriores y podría tratarse de un linaje paralelo al de los homininos posterior a la divergencia del chimpancé.
- Brunet M, Guy F, Pilbeam D, et al. (2002) A new hominid from the Upper Miocene of Chad, Central Africa. Nature 418, 145–151.
- Brunet M, Guy F, Pilbeam D, et al. (2005) New material of the earliest hominid from the Upper Miocene of Chad. Nature 434, 752–755.
- Guy F, Lieberman DE, Pilbeam DR, et al. (2005) Morphological affinities of the Sahelanthropus tchadensis (Late Miocene hominid from Chad) cranium. Proc Natl Acad Sci USA 102, 18836–18841.
- Zollikofer C, de Leon M, Lieberman DE, et al. (2005) Virtual cranial reconstruction of Sahelanthropus tchadensis. Nature 434, 755–759.
- Lebatard AE, Bourle’s DL, Duringer P, et al. (2008) Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. Proc Natl Acad Sci USA 105, 3226–3231.
- Emonet, E-G, Andossa, L, Mackaye, HS, et al. (2014) Subocclusal dental morphology of Sahelanthropus tchadensis and the evolution of teeth in hominins. Am J Phys Anthropol 153, 116–123.
Orrorin tugenensis
Tugen Hills, Kenia (6,1-5,7 ma)

Orrorin tugenensis
Esta especie del Mioceno fianl de Tugen Hills en Kenia está datada en 6,1-5,7 ma. Los restos encontrados incluyen dos fragmentos proximales de femur izquierdos (BAR 100200, BAR 100300), otro derecho (BAR 121500), un fragmento izquierdo de mandíbula (BAR 100000), un fragmento distal de húmero (BAR 100400) y algunos dientes aislados. El análisis de la morfología del frangmento proximal del femur de Orrorin muestra más similitudes con la morfología de los homininos bípedos que con los con los hominoideos actuales, cuya morfología sería más derivada debido a adaptaciones para la la braquiación suspensora. Las diferencias anatómicas entre Orrorin y los australopitecinos en general sugieren que Orrorin tenía un bipedismo diferente al de los humanos modernos y más parecido al de Ardipithecus ramidus, con una tuberosidad isquiática lateralizada, debajo del trocanter mayor, y pequeña (parta la inserción del múscuylo gluteo máximo), y una incipiente línea áspera anchar. Por tanto, la configuración anatómica de los humanos modernos (inserción posterior del gluteo máscimo) no estaba presente en los homininos más primitivos. Para este caracter, Orrorin es intermedio entre los hominoideos del Mioceno y los australopitecinos.

Posición filogenética de Orrorin tugenensis
En base a la morfología dental y postcraneal se ha sugerido que Orrorin pertenece al linaje de los homininos. Los molares son más pequeños que los de los australopitecinos y parecidos en tamaño a los de Ardipithecus. La dentición anterior, incisivos y caninos superiores, así como el P4 inferior, en cambio, difieren de los de los homininos posteriores y se parecen más a los del chimpancé, pero los molares tienen el esmalte grueso. Orrorin tiene dientes yugales pequeños respecto al tamaño corporal (rasgo primitivo heredado del ancestro común de todos los hominoideos africanos) presente también en Homo). Se ha sugerido que Orrorin tugenensis ya estaba adaptado a un bipedismo habitual en el suelo, incluso obligado, aunque también era un buen escalador arborícola. El femur y el húmero de Orrorin son 1,5 veces mayores que los de AL 288.1 (A. afarensis), con una talla similar a la de una hembra de chimpancé actual. La posición filogenética de esta especie es problemática: tiene un bipedismo derivado que sugiere que no sería el ancestro de Ardipithecus, que tiene una condición más primitiva, pero la cabeza del femur de Orrorin es intermedia entre la de los hominoideos del Mioceno y los homininos posteriores.
- Senut B, Pickford M, Gommery D, et al. (2001) First hominid from the Miocene (Lukeino Formation, Kenya). C R Acad Sci Paris 332, 137–144.
- Pickford M, Senut B, Gommery D, Treil J (2002) Bipedalism in Orrorin tugenensis revealed by its femora. C. R. Palevol. 1, 191–203.
- Galik K et al. (2004) External and internal morphology of the BAR 1002’00 Orrorin tugenensis femur. Science 305, 1450–1453.
- Richmond BG, Jungers WL (2008) Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science 319, 1662–1665.
- Almécija S, Tallman M, Alba DM, et al. (2013) The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nat Comm 4, 2888.
Ardipithecus kadaba
Gona, Etiopía (6,3-5,4 ma)

Comparación del premolar ABD1/P1 (arriba) con ALA VP-2/10 (abajo) de Ardipithecus kadaba.
Los restos fósiles (5 dientes) más antiguos atribuidos a la especie Ardipithecus kadaba estan datados en 5,4 ma y 6,3 ma en la formación Adu-Asa en el área de Gona en Etiopía. Datos sedimentológicos e isotópicos sugieren un hábitat de selvas o bosques hebaceos en una zona con abundantes cursos de agua y lagos. Estos dientes (LRP4, URC, ULM3, LLP3, ULM1/2) no presentan apomorfías que lo aproximen a los hominoideos no humanos,. Al contrario, muestra rasgos derivados en la dirección de los homininos posteriores, en particular por la reducción o ausencia del patrón de desgaste sectorial del complejo C/P3.
Simpson SW, Kleinsasser L, QuadeJ, McIntosh WC, Dunbar N, Semaw S, Rogers MJ (2015) Late Miocene hominin teeth from the Gona Paleoanthropological Research Project area, Afar, Ethiopia. J Hum Evol 81, 68–82.
Middle Awash, Ethiopia (5,8-5,2 ma)
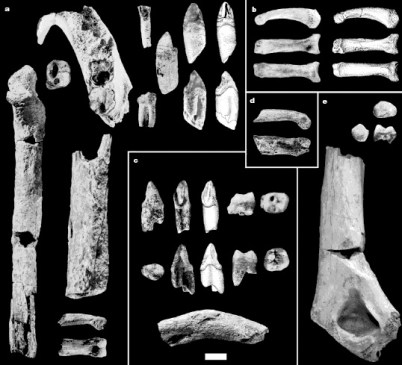
A. kadaba de Middle Awash: a) ALA VP-2/10 mandíbula y dientes, b) AME VP-1/71 falange del pie, c) STD VP-2 dientes y clavícula, d) DID VP-1/80 falange de la mano, e) ASK VP-3/160 diete, ASK VP-3/78 fragmento distal de húmero.
Los fósiles de los yacimientos del Mioceno fianl de Middle Awash en Etiopía se asignaron en 2001 a la subespecie Ardipithecus ramidus kadabba. Posteriormente, en noviembre de 2002 nuevos fósiles sirvieron para definir la especie A. kadabba. Se encontraron 6 dienteslar en la localidad 3 de Asa Koma (ASK-VP-3), datada en 5,6 a 5,8 ma. Los caracteres derivados de estos dientes son compartidos con todos los los homininos posteriores, lo que indica que son posteriores a la divergencia de loslinajes panini y hominini. Sin embargo, la persistencia de algunos caracteres primitivos de la dentición y del esqueleto postcraneal en estos fósiles indica que Ardipithecus kadabba es un taxón filogeneticamente próximo al ancestro común de chimpancés y humanos. Los dientes de Ardipithecus kadabba muestran un mosaico de características. El esmalte dental de los homininos de las localidades STD y ALA son parecidos, aunque algo mayores, a los de Ardipithecus ramidus de Aramis. La presencia de 4 cúspides diferenciadas y la ausencia de afinamiento distal del M3 son rasgos primitivos compartidos con los hominoideos del Mioceno. El desarrollo del tubérculo dental en los caninos inferiores y la posición variable del ‘hombro’ mesial de la corona y la expansión de la cresta marginal mesial son rasgos derivados. La diáfisis humeral es pequeña y se parece a la del menor de los Australopithecus afarensis. Aunque incompleta, la diáfisis de la ulna está más curvada que en los homininos posteriores.
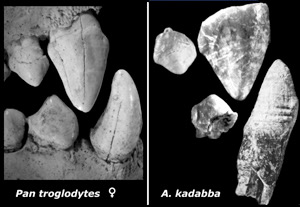
Complejo cortador C/P3 del chimpancé (izquierda) y caninos superior (ASK VP-3/400) e inferior (STD VP-2/61), premolar superior (ASK VP-3/160) e inferior (ASK VP-3/403) de A. kadaba.
La morfología primitiva de los fósiles del Mioceno de Middle Awash sugieren que A. kadabba es distinto a A. ramidus y que en ancestro común del chimpancé y los humanos tenía un complejo C/P3 funcional. La comparación con los dientes de Sahelanthropus y Orrorin suggests con A. kadabba muestra muchas similitudes. El P3 inferior de A. ramidus, Au. afarensis o Au. anamensis carecen de faceta de contascto en la cara bucal de la corona, orientada mesio-bucalmente, En cambio, el complejo C/P3 está presente en el material de Asa Koma, por lo que A. kadabba se ha descrito como una especie separada de A. ramidus. El único canino superior conservado en Orrorin no está suficientemente desgastado para determinar si presenta el borde distal del complejo cortador. Los pocos elementos conservados del complejo C/P3 en S. tchadensis parece que no muestran señales de este tipo de desgaste. Los datos métricos y anatómicos del material conservado de A. kadabba, O. tugenensis y S. tchadensis no son suficientes para mantener su separación taxonómica, dejando aparte las diferencias en el grosor del esmalte entre ellas, y es posible que se puedan agrupar en un solo género.
- Haile-Selassie Y (2001) Late Miocene hominids from the Middle Awash, Ethiopia. Nature 412(6843), 178-181.
- WoldeGabriel G, Haile-Selassie Y, Renne PR, Hart WK, Ambrose MH, Asfaw B, Heiken G, White TD (2001) Geology and paleontology of the Late Miocene Awash valley, Afar Rift, Ethiopia. Nature 412(6843), 175-178.
- Haile-Selassie Y, Suwa G, White TD (2004) Late Miocene teeth from the Middle Awash, Ethiopia, and early hominid dental evolution. Science 303(5663), 1503–1505
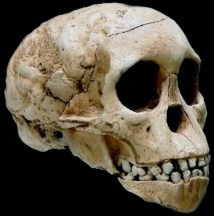
Ardipithecus ramidus
Middle Awash, Etiopía (4,4 ma)
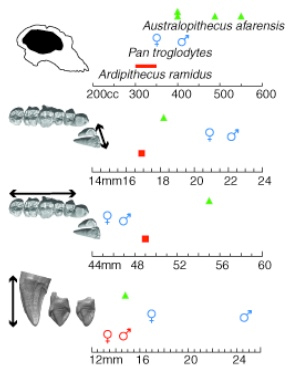
Comparación de la capacidad craneal y tamaño dental de Ar. ramidus, Au. afarensis y Pan.
En octubre de 2009 la revista Science publica una monografía sobre un nuevo taxón encontrado en Middle Awash entre 1992 y 1995, Ardipithecus ramidus, inicialmente atribuido al género Australopithecus. Additional fossils of the fairly complete specimen appeared in 1995 and following field seasons, that were finally adscribed to a distinct genus: Ardipithecus. Sus rasgos anatómicos, especialmente del especimen denominado ‘Ardi’, le diferencian claramente de Australopithecus afarensis. El especimen ARA-VP-6/500 incluye la mayor parte de la dentición y la cara, calota craneal y base de craneo. Su morfología craneofacial muestra una reducida capacidad craneal (300-350 cc), una reducida proyección mediofacial y ausencia del prognatismo subnasal característico de los hominoideos no humanos. La base de su craneo es corta, lo que difiere de Pan troglodytes y P. paniscus. Ardipithecus ramidus no tiene el desarrollo tan marcado del esqueleto zygomaxilar de Australopithecus. Esta combinación de caracteres parece que es compartida con Sahelanthropus, lo que sugiere que el craneo de los homininos de Mioceno-Plioceno es distinto al de los hominoideos actuales y también del de Australopithecus.
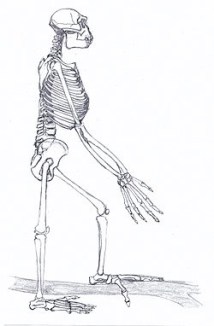
Esqueleto de Ardipithecus ramidus.
La cara de Ar. ramidus es corta y gracil, con un prognatismo facial parecido al de Sahelanthropus, y su craneo y el torus supraorbitario difieren de los del chimpancé. Ardipithecus ramidus tiene los incisivos pequeños, comparado con el chimpancé, pero la dentción yugal es algo mayor (aunque es menor que en Australopithecus) y los caninos son pequeños y no presenta dimorfismo sexual, lo que sugiere grupos sociales menos agresivos. El pie muestra un dedo gordo claramente divergente, lo que junto con otras evidencias esquelléticas sugiere que el último ancestro común de humanos y chimpancés no tenía ni la mano, el pie, la pelvis, vértebras o extremidades adaptadas a la locomoción suspensora, trepadora o cuadrumana de los hominoideos africanos actuales. La anatomía del esqueleto de chimpancés y gorilas parecen ser modelos poco apropiados para explicar el origen y evolución de bipedismo. Las adaptaciones locomotoras (apomorfías) de Ar. ramidus sugieren hábitos arboreos de escalados palmígrado con adaptaciones al bipedismo terrestre más primitivas que Australopithecus. El complejo C/P3 de Ar. ramidus era reducido y análisis isotópicos sugieren una dieta basada en el consumo de plantas C3 en un hábitat de bosque cerrado. Es probable que el LCA entre chimpancés y humanos haya sido distinto a todos los hominoideos africanos actuales.
- White TD, et al (2009) Ardipithecus ramidus and the paleobiology of early hominids. Science 326, 64–86.
- WoldeGabriel G, et al (2009) The geological, isotopic, botanical, invertebrate, and lower vertebrate surroundings of Ardipithecus ramidus. Science 326, 65-65e5.
- Louchart A, et al (2009) Taphonomic, avian, and small-vertebrate indicators of Ardipithecus ramidus habitat. Science 326, 66–66e4.
- White TD et al (2009) Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 67–93.
- Suwa G et al. (2009) The Ardipithecus ramidus skull and its implications for hominid origins. Science 326, 68–68e7.
- Suwa G et al. (2009) Paleobiological implications of the Ardipithecus ramidus dentition. Science 326, 69–99.
- Lovejoy CO et al (2009) Careful climbing in the Miocene: the forelimbs of Ardipithecus ramidus and humans are primitive. Science 326, 70–70e8.
- Lovejoy CO et al (2009) The pelvis and femur of Ardipithecus ramidus: The emergence of upright walking. Science 326, 71–71e6.
- Lovejoy CO et al (2009) Combining prehension and propulsion: the foot of Ardipithecus ramidus. Science 326, 72–72e8.
- Lovejoy CO (2009) The great divides: Ardipithecus ramidus reveals the postcrania of our last common ancestors with African Apes. Science 326, 73–106.
- Lovejoy CO (2009) Reexamining human origins in light of Ardipithecus ramidus. Science 326: 74–74e8.
Lothagam, Kenya (4,2 ma) y Tabarin, Kenia (4,15 ma)

Mandíbulas de Tabarin (izquierda) y Lothagam (derecha)
Los fósiles del Mioceno final de Lothagam (4,2 ma) y Tabarin (Baringo, 4,15 ma) han sido atribuidos también el género Ardipithecus. Los fragmemtos mamdibulares de Lothagam (KNM-LT 329) y Tabarin (KNM-TH 13150) aportan poco información sobre su origen. Lothagam se parece a Au. afarensis por laposición de foramen mentoniano, la anchura del margen alveolar, el origen de la rama mandibular, la reducción del hypoconúlido y el tamaño del primer molar. Pero la presenta algunas rasgos de los hominoideos, como la apertura del foramen mentoniano y la presencia de refuerzo sinfisario (simian shelf)cuestionan su afinidad con los homininos.
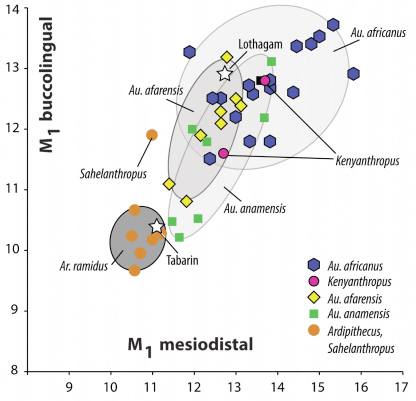
Diámetros mesio-distal y buco-lingual del primer molar inferior de Tabarin y Lothafam.
El tamañoa del molar está en el rango de Au. afarensis, pero el resto de rasgos morfológicos son primitivos y coinciden con los de diversos taxones. Sim embargo, las mandíbulas de Sahelanthropus, Ar. ramidus, Lothagam, y Tabarin muestran similitudes morfológicasare con Ar. kadabba y difieren de Au. anamensis y Au. afarensis ya que son más pequeñas y gráciles, y tiene el borde lateral de la rama más ligero, por lo que ambas mandíbulas son más parecidas a Ardipithecus que a Australopithecus. El análisis métrico de la dentición de Tabarin también sugiere su atribución a Ardipithecus, lo que ampliaría el rango geográfico de estya especie. Lothagam podría no encajar tan bien en esta especie, ya que el tamaño del molar es bastante grande y se parece más en esto a Au. afarensis y Au. anamensis. En este sentido, la datación de Lothagam es crucial para determinar si Ardipithecus y Australopithecus coexistieron o no en la misma cronología. La clasificación de Tabarin en Ar. ramidus parece más clara .
- Haile-Selassie Y (2001) Late Miocene hominind from the Middle Awash, Ethiopia. Nature 412, 178–181.
- WoldeGabriel G, Haile-Selassie Y, Renne PR, et al (2001) Geology and palaeontology of the Late Miocene Middle Awash valley, Afar rift, Ethiopia, Nature 412(6843), 175-178.
- Leakey MG, Walker AC (2003) The Lothagam hominids. In: Leakey MG, Harris JM (Eds.) Lothagam: The dawn of humanity in Eastern Africa. Columbia University Press, New York, pp. 249-257.
- Haile-Selassie Y, Suwa G, White TD (2004) Late Miocene teeth from Middle Awash, Ethiopia, and early hominid dental evolution. Science 303, 1503–1505.
- Simpson SW, Kleinsasser L, Quae J, et al. (2015) Late Miocene hominin teeth from the Gona Paleoanthropological Research Project area, Afar, Ethiopia. J Hum Evol 81, 68–82.
- Kissel M, Hawks J (2015) What are the Lothagam and Tabarin Mandibles? PaleoAnthropology 2015: 37−43.
Australopithecus anamensis
Los primeros restos atribuidos a Australopithecus anamensis se encontraron en Kanapoi, en Kenia. Otros restos atribuidos a esta especie proceden también de Allia Bay (Kenia), Asa Issie (Etiopía) y Woranso-Mille (Afar, Ethiopia).
Kanapoi, Kenia (4,2-4,1 ma)
Restos dentales, craneales y postcraneales de nueve especímenes proceden de Kanapoi, Kenya. Un mosaico de rasgos primitivos y derivados los situan como ancestro de Australopithecus afarensis, mientras que Ardipithecus ramidus podría ser un taxón hermano (sister species). La tibuia, que indica que ya era bípedo, el humero y la mandíbula (KNM-KP 29287, probablemente de un macho por su gran tamaño) son algo más reciente que los otros materiales del mismo yacimiento. Las dataciones confirman fechas intermedias entre Ar. ramidus and Au. afarensis. Los materiales encontrados demuestran que es más primitivo que Au. afarensis. Dataciones de 40Ar-39Ar indican que, excepto una mandíbula, todos los materiales de Kanapoi proceden de sedimentos depositados entre 4,17 y 4,07 ma. Las excavaciones en Kanapoi entre 2003 y 2008 proporcionaron 9 nuevos fósilesdatados entre 4,195 y 4,108 ma. Estos materiales incluyen un fragmento de mandíbula sin dientes, un fragmento de maxilar, dientes en un fragmento mandibular y otros aislados. Todos los homininos de Kanapoi comparten una morfología común del complejo C/P3, con el canino bajo pero con una cresta mesiodistal del P3 más larga que en Au. afarensis, y raíces del canino largas (probablemente dimorficas), que no varían respecto a la condición primitiva. El paso de Au. anamensis a Au. afarensis comportó cambios muy significativos en la dentición anterior y en la estructura del maxilar y los dientes molares, lo que podría estar relacioado con un cambio de dieta. La corona del canino no cambia, pero el P3 superior se acorta mesiodistalmente, la corona se hace más simétrica, y los P3s se vuelven menos monocúspides.
Allia Bay, Kenia, 3,95 ma)
El material de Allia Bay es similar al de Kanapoi y se ha clasificado en la misma especie, que habría sido un bípedo habitual, aunque retiene rasgos primitivos en los brazos. El Complejo C/P3 es algo más primitivo que el que se observa en in Au. afarensis, pero no tanto como el de Ar. ramidus. El material de Allia Bay conserva menos caracteres de diagnóstico queel de Kanapoi, pero su similidud y proximidad geográfica permiten clasificarlos juntos. Australopitheus anamensis tenóa el esmalte grueso en los molares y la dentición anterior era pequeña, los caninos mandibulares eran estrechos y asimétricos. El esmalte grueso y los incisivos pequeños sugieren una dieta abrasiva como resultado de un cambio a largo plazo desde un hábitat cerrado hacia hábitats más abiertos.
Asa Issie, Etiopía (4,2–4,1 ma)
Estos fósiles de la región de Middle Awash amplían el rango de distribución de Au. anamensis al norte de Etiopía. Proceden de una región estratigraficamente controlada y datada en 4,1-4,2 ma. Los materiales incluyen restos craneales y dentales, con un canino de gran tamaño y el femur más antiguo de Australopithecus. La reconstrucción ecológica es un bosque cerrado. La cronología y los rasgos anatómicos intermedios de estos materiales están entre Ar. ramidus and Au. afarensis, lo que sugiere un cambio evolutivo rápido de Ardipithecus a Australopithecus en esta región de África, ya sea por sustitución poblacional o por una evolución filogenética acelerada.
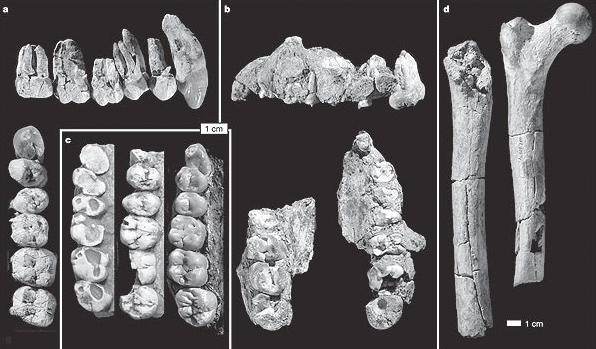
Asa Issie fossil remains of Au. anamensis. a) ASI VP-2/334 dentición superior derecha, b) ARA VP-14/1 maxilar superior con dientes, c) KNM KP-2928 (izquierda), KNM ER-30745 (centro), AL-200-1 Au. afarensis (derecha), d) ASI VP5/154 diáfisis de femur derecho (izquierda) y AL 288-1 Lucy Au. afarensis (derecha).
Woranso-Mille, Ethiopia (3.8–3.57 ma)
Estos materiales de Woranso-Mille están datados en 3,57–3,8 ma. En su mayoría se trata de dientes aislados, fragmentos mandibulares y maxilares, y algunos fragmentos postcraneales. La morfología dental muestra mayor afinidad con Au. anamensis de Allia Bay/Kanapoi (Kenia) and Asa Issie (Etiopía) que con Au. afarensis de Hadar (Etiopía). Sin embargo, son intermedios en la morfología dental y mandibular, en particular al compararlos con el material de Laetoli. Kanapoi, Allia Bay y Asa Issie representan la condición primitiva de Au. anamensis de Hadar, mientras qu Laetoli yWoranso-Mille representan un mosaico.
Los materiales de Woranso-Mille son particularmente interesantes porque cubren el rango temporal entre 3,5 y 3,8 ma y sugieren que podría tratarse de cronoespecies. Dentalmente los materiales de Woranso-Mille son más parecidos a Au. anamensis de Allia Bay que a Au. afarensis de Laetoli, pero tienen algunos caracteres derivados compartidos con Au. afarensis de Hadar y Laetoli.
- Leakey MG, Feibel CS, McDougall I, Walker A (1995) New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature 376, 565–571.
- Leakey MG, Feibel CS, McDougall I,Ward CV, Walker A (1998) New specimens and confirmation of an early age for Australopithecus anamensis. Nature 393, 62–66.
- Ward CV, Leakey MG, Walker A (2001) Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. J Hum Evol 41(4), 255-368. Erratum in: J Hum Evol 2001 41(6), 713–716.
- White TD, WoldeGabriel G, Asfaw B, et al (2006) Asa Issie, Aramis and the origin of Australopithecus. Nature 440(7086), 883-889.
- Haile-Selassie Y, Saylor BZ, Deino A et al (2010) New hominid fossils from Woranso-Mille (Central Afar, Ethiopia) and taxonomy of early Australopithecus. Am J Phys Anthropol 141, 406–417.
- Ward CV, Plavcan JM, Manthi FK (2010) Anterior dental evolution in the Australopithecus anamensis–afarensis lineage. Phil Trans R Soc B 365, 3333–3344.
- Ward CV, Manthi FK, Plavcan JM (2013) New fossils of Australopithecus anamensis from Kanapoi, West Turkana, Kenya (2003-2008) J Hum Evol 65, 501–524.
Australopithecus deyiremeda
Woranso-Mille, Ethiopia (3,5–3,3 ma)
La diversidad de los homininos del Pleistoceno Medio está sujeta a importantes debates, especielmente respecto a los taxones Australopithecus bahrelghazali y Kenyanthropus platyops en comparación con Australopithecus afarensis. A estas especies se añaden los fósiles descritos por Haile-Selassie et al. (Nature 521, 483–488) y atribuidos a una nueva especie: Australopithecus deyiremeda datada en 3,3–3,5 ma en los depósitos de Woranso–Mille de la región central de Afar en Etiopía. La morfología facial y dental de esta especie sugiere un cierto grado de convergencia (homoplasia) con Paranthropus y Homo en un periodo anterior al que se pensaba.
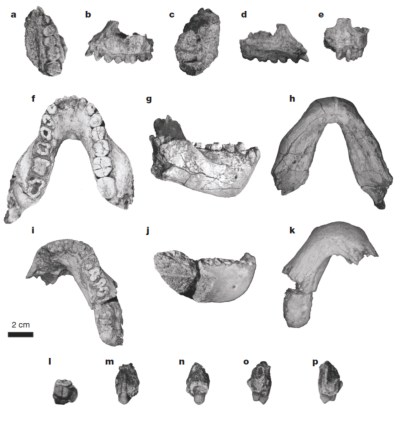
Holotipo BRT-VP-3/1. a, vista oclusal. b, lateral. c, superior. d. medial. e, anterior. Paratipo BRT-VP-3/14. f, vista oclusal. g, lateral derecha. h, basal. i, oclusal. j, lateral derecha. k, basal. Especiemn BRT-VP-3/37. 1, vista oclusal. m, buccal. n, lingual. o, distal. p, mesial.
La morfología de las mandíbulas BRT-VP-3/14 y WYT-VP-2/10 es parecida a la de las formas robustas de África del Este, aunque su tamaño es parecido al de Au. afarensis. El origen de la rama ascendente se sitúa en el P4 como en la mandíbula de Peninj (Paranthropus boisei) y SK-12 (P. robustus). En Australopithecus afarensis el inicio de la rama mandibular se sitúa más atrás, a nivel de los molares. BRT-VP-3/14 y WYT-VP-2/10 tienen cuerpos mandibulare robustos, especialmente a nivel de M2–3, y las dimensiones de BRT-VP-3/14 y WYT-VP-2/10 en esta zona son considerables. Es decir, son muy anchas para su reducido tamaño. Además, la mandíbula LD 350-1 del yacimiento de Ledi–Geraru datada en 2,8 ma y atribuida a Homo, se parece también a las de Au. deyiremeda y en alguna medida también a las de Au. afarensis en la morfología del cuerpo mandibular. Pero los caracteres derivados de LD 350-1, característicos de Homo pero ausentes en Au. deyiremeda (como la posición retrasada del foramen mentoniano y el M3 más corto en sentido mesiodistal que el M2).
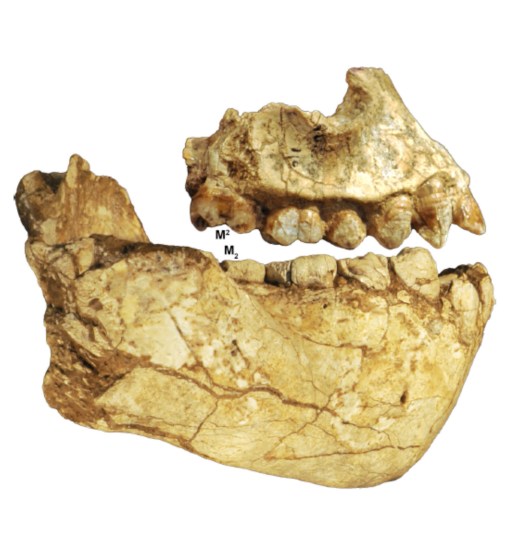
BRT-VP-3/1 (girado) y BRT-VP-3/14
El análisis filogenético de Au. deyiremeda sugiere que se debe incluir en el clado formado por Au. africanus, Paranthropus y Homo, pero sus diferencias con estos dos géneros sugiere que las similitudes con ellos podrían haber aparecido independientemente. Woranso–Mille está geograficamente muy próximo a Hadar (35 Km al norte), donde Au. afarensis está documentado. Sin embargo, las diferencias con este taxón podrían indicar la presencia de diversas especies de homininos en el mismo momento cronológico y en zonas muy próximas. La comparación del holotipo de Kenyanthropus platyops KNM-WT 40000 y el holotipo de Australopithecus deyiremeda BRT-VP-3/1 con Australopithecus afarensis muestran que tanto KNM-WT 40000 como BRT-VP-3/1 difieren en la forma del maxilar superior de A. afarensis, pero de formas distintas, siendo Au. deriyemeda la que menos se parece a A. afarensis. Este análisis sugiere que los dos especímenes deben mantenerse en una especie separa da de A. afarensis.
- Spoor F, Leakey MG, O’Higgins P. (2016) Middle Pliocene hominin diversity: Australopithecus deyiremeda and Kenyanthropus platyops. Philos Trans R Soc Lond B Biol Sci 371(1698) pii, 20150231.
Australopithecus afarensis
Hadar, Ethiopia (3,4-2,96 ma)
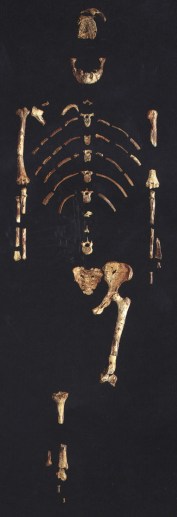
AL 388-1 Lucy Au. afarensis
De Hadar, Etiopía 3,4-2,96 ma (Miembros Sidi Hakoma, Denen Dora y Kada) proceden unps 367 especímenes (90% del total de la especie); 57 mandíbulas de adulto o subadulto y 12 fragmentos craneales. Se trata de uno de los taxones mejor conocidos del registro fosil. La mayoría de restos fósiles proceden de la localidad de Afar (Formación Hadar). Destaca el esqueleto parcial AL 288-1 (Lucy de 3,3 ma) atribuido a la especei Australopithecus afarensis cuyo esqueleto difiere del chimpancé y el craneo es parecido al de Australopithecus africanus. Su anatomía sugiere una conducta suspensora a la vez que bípeda (bipedismo facultativo), con un cerebro pequeño pero con un mayor desarrollo y complejidad respecto al del chimpancé.
Se trata de uno de los taxones mejor conocidos del registro fosil. La mayoría de restos fósiles proceden de la localidad de Afar (Formación Hadar). Destaca el esqueleto parcial AL 288-1 (Lucy de 3,3 ma) atribuido a la especei Australopithecus afarensis cuyo esqueleto difiere del chimpancé y el craneo es parecido al de Australopithecus africanus. Su anatomía sugiere una conducta suspensora a la vez que bípeda (bipedismo facultativo), con un cerebro pequeño pero con un mayor desarrollo y complejidad respecto al del chimpancé.
- AL 129-1 (Donald Johanson, 1973). 3,4 ma. Porciones de ambas piernas, incluyendo una rodilla derecha pequeña de un individuo adulto.
- AL 288-1, Lucy (Donald Johanson y Tom Gray, 1974). 3,2 ma. Hembra adulta de 25 años (40% del esqueleto). La pelvis, fémur y tibia indican que era un bípedo de 107 cm de estature y 28 Kg.
- AL 333 Site, The First Family (Donald Johanson, 1975). 3,2 ma. Restos pertenecientes a 13 individuos.
- AL 333-1. Mandibula derecha infantil.
- AL 333-105. Cráneo juvenil parcial.
- AL 333-160. Cuarto metatarsiano izquierdo (Ward, Kimbel y Johanson, 2011).
- Abundantes restos de miembros superiores e inferiores.
- AL 400-1a, mandíbula halladan entre 1974-1977 por Don Johanson. Está datada en 3 ma. Relativamente larga y estrecha, con molares de tamaño moderado con patrón Y5 e incisivos relativamente anchos.
- AL 417-1. Cráneo parcial.
- AL 438-1. Esqueleto parcial con asociaciones craneodentales.
- AL 444-2. (Bill Kimbel y Yoel Rak, 1991). 3 ma. Hallado por Tim White. Cráneo con el 70% de los huesos, de un macho adulto con capacidad craneal de 550 cc. De la comparación con Lucy se deduce un gran dimorfismo sexual.
- AL 822-1. Cráneo casi completo hallado en 2000. Probablemente femenino por su pequeño tamaño.
- AL 1030-1. (Halszka Glowacka, William H Kimbel y Donald C Johanson, 2017). Fragmento del corpus mandibular izquierdo de un infante con dp3 y dp4 erupcionados. Son visibles las coronas de I1 e I2 no erupcionadas. Conserva la raíz del ramus ascendente. Recuperado en 2002. Datado en 3,2 ma.
- AL 1920-1. (Halszka Glowacka, William H. Kimbel y Donald C. Johanson, 2017). Mandíbula paracial de un juvenil que conserva completo el corpus derecho con el nacimiento del ramus ascendente y la región sinfisal con el corpus anterior izquierdo adyacente. Recuperado en 2012, con una datación de ca 3,35 ma.
Maka, Etiopía (3,8-2,9 ma)
Desde su descrubimiento el debate se centró en si los materiales de Kenia y Tanzania atribuidos a A. afarensis representan especies distintas o si la variabilidad observada es atribuible a dimorfismo sexual. Los fósiles descubiertos en la región de Afar en Etiopíahiopia en 1990 fueron durante la década de los 70 del siglo XX los homininos más antiguos conocidos. Nuevos hallazgos procedentes de Maka (algunas mandíbulas, una de ellas casi completa, fragmento proximal de un fémur y un húmero), datados en 3,4 ma sugieren que todos los materiales pertenecen a una única especie, aunque adaptada a ambientes diversos, sexualmente dimórfica y bipeda. Los fósiles de Maka muestran una gran variabilidad de tamaños. La casi completa mandíbula MAK-VP-1/12 muestra muestra grandes similitudes dentales con los materiales de Hadar y Laetoli y reflejan la transición de A. anamensis a A. afarensis en lo que respecta a la progresiva megadoncia relacionada con el consumo de alimentos más abrasivos.
Worason-Mille, Etiopía (3,33-3,20 ma)
Especímenes fósiles de Au. afarensis han aparecido en el yacimiento de Nefuraytu (datado en 3,330-3,207 ma (Woranso-Mille, central Afar, Etiopía; Haile-Selassie et al, 2016). El material incluye el especimen NFR-VP-1/29, una mandíbula casi completa de gran tamaño, probablemente de un infividui masculino, que muestra todos los caracteres distintivos de Au. afarensis (simian shelf y foramen mentoniano avanzado, entre otros). Otros materiales incluyen: NFR-VP-1/2 (corona sin erupcionar de LRM1), NFR-VP-1/58 (LRM2/3), NFR-VP-1/96 (fragmento de canino inferior), NFR-VP-1/213 (URM3) y NFR-VP-1/214 (fragmento de hueso frontal). También detaca KSD-VP-1/1 Kadanuumuu, que incluye fragmentos postcraneales (Haile-Selassie et al, 2010; Yohannes Haile-Selassie, Denise F Su, 2015) de un macho adulto hallado en Woranso-Mille 40 km al norte de Hadar, Gona y Dikika, y datado en hace 3,6 Ma (Saylor et al, 2015). El material incluye 6 gragmentos de vértebras cervicales (C2-C7) más derivadas que las de Lucy y Selam (Meyer, 2015).Kadanuumuu muestra la única tibia completa conservada de Au. afarensis.Su tórax revela una postura completamente erguida (Latimer et al, 2015).
Dikika, Etiopía (3,3 ma)
DIK-1/1, Selam, Bebé Dikika (Zeresenay Alemseged, 2000). Es un cráneo y esqueleto bastante completo de una niña de 3 años. El volumen craneal corresponde a un 65-88% del típico de la especie. En los chimpancés a los tres años se alcanza el 90%.En la localidad DIK-55 fueron hallados dos huesos con marcas de corte, datados en 3,39 ma (McPherron et al 2010; Thompson et al 2015).
East Turkana, Kenia (3,35 ma)
El fragmento de mandíbula y varios dientes aislados procedentes de sedimentos fluviales datados en más de 4 ma procedentes de East Lake Turkana recuerdan a los materiales de Laetoli (Tanzania) y Hadar (Etiopía) asignados a Australopithecus afarensis. Además, fósiles del Lago Turkana (KNM-ER 2602, fragmento de calota craneal), datadosen 3,35 a 1,0 ma, situados por encima de Tulu Bor Tuff (Koobi Fora) en el miembro Lomekwi de la formación Nachukui se han atribuido también a Australopithecus afarensis. Esta especie es rara en Kenia, probablemente por la escasez de sedimentos de este periodo..
Laetoli, Tanzania (3,7-3,5 ma)
El yacimiento de Laetoli está situado cerca del de Olduvai Gorge al norte de Tanzania. Fue excavado en los años 1930 por Louis y Mary Leakey, pero entonces no se encontraron fósiles. Mary Leakey regresó a Laetoli con Tim White en 1978 y encontraron huesos atribuibles a Australopithecus afarensis datatos en 3,7-3,5 ma (más antiguos que Lucy). Entre los restos más destacados están Garusi I (mandíbula),LH-2 (mandíbula),LH-4 (holotipo),LH-5 (mandíbula) y LH-21 (esqueleto juvenil parcial).
También encontraron 59 huellas de homininos bípedos sobre un sustrato de cenizas volcánicas. Las huellas corresponden a dos individuos que caminaron juntos hace unos 3,5 ma y muestran un dedo gordo alineado con el resto de dedos, no divergente.
- Johanson DC, White TD (1979) A systematic assessment of early African hominids. Science 203(4378), 321-330.
- White TD (1980) Additional fossil hominids from Laetoli, Tanzania: 1976–1979 specimens. Am J Phys Anthropol 53(4), 487-504.
- Johanson DC, et al (1982) Morphology of the Pliocene partial hominid skeleton (AL 288‐1) from the Hadar formation, Ethiopia. Am J Phys Anthropol 57(4), 403-451.
Schmid P (1983) A reconstruction of the skeleton of A.L. 288-1 (Hadar) and its consequences. Folia Primatol40(4), 283-306. [artículo en alemán]
- Stern Jr JT, Randall LS (1983) The locomotor anatomy of Australopithecus afarensis. Am J Phys Anthropol 60(3), 279-317.
Holloway RL (1983) Cerebral brain endocast pattern of Australopithecus afarensis hominid. Nature303(5916), 420-422.
Kimbel WH, White TD, Johanson DC (1984) Cranial morphology of Australopithecus afarensis: a comparative study based on a composite reconstruction of the adult skull. Am J Phys Anthropol64(4), 337-388.
- White, Tim D., and Gen Suwa (1987) Hominid footprints at Laetoli: facts and interpretations. Am J Phys Anthropol 72(4), 485-514.
Kimbel WH, Johanson DC, Rak Y (1994) The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia. Nature368(6470), 449-451.
White TD, Suwa G, Simpson S, Asfaw B (2000) Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia. Am J Phys Anthropol111(1), 45-68.
- Lockwood CA, Kimbel WH, Johanson DC (2000) Temporal trends and metric variation in the mandibles and dentition of Australopithecus afarensis. J Hum Evol39(1), 23-55.
Brown B1, Brown FH, Walker A (2001) New hominids from the Lake Turkana Basin, Kenya. J Hum Evol41(1), 29-44.
- Holloway RL, Yuan MS (2004) Endocranial Morphology of A. L. 444-2. In WH Kimbel, Y Rak, DC Johanson (Eds.) The skull of Australopithecus afarensis. New York: Oxford University Press, pp. 123-135.
- Kimbel, W. H., Rak, Y. & Johanson, D. (2004). The skull of Australopithecus afarensis. New York: Oxford University Press.
Alemseged Z, et al (2005) A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context. Hum Evol49(4), 499-514.
- Alemseged Z, et al (2006) A juvenile early hominin skeleton from Dikika, Ethiopia. Nature 443(7109), 296-301.
- Haile-Selassie Y, Latimer BM, Alene M, Deino AL, Gibert L, Melillo SM, Saylor BZ, Scott GR, Lovejoy CO (2010) An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proc Natl Acad Sci USA107(27), 12121-12126.
- McPherron SP, et al (2010) Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature 466(7308), 857-860.
- Raichlen DA et al (2010) Laetoli footprints preserve earliest direct evidence of human-like bipedal biomechanics. PLoS One 5(3), e9769.
- Ward CV, Kimbel WH, Harmon EH, Johanson DC (2012) New postcranial fossils of Australopithecus afarensis from Hadar, Ethiopia (1990-2007). J Hum Evol 63(1), 1-51.
Haile-Selassie Y et al (2016) Dentognathic remains of Australopithecus afarensis from Nefuraytu (Woranso-Mille, Ethiopia): Comparative description, geology, and paleoecological context. J Hum Evol100:35-53.
Australopithecus bahrelghazali
Bhar el Ghazal, Chad (3,4-3 ma)
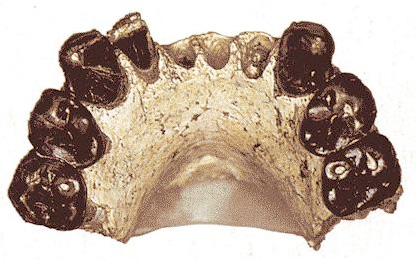
KT 12/H1 Abel.
Esta especie se definió a partir del hallazgo en 1995 de un fragmento mandibular (holotipo: KT 12/H1, Abel) con 7 dientes (1 I, 2 P y 2 C) atribuidos a una nueva especie Au. bahrelghazali, encontrado en Koro Toro, Desierto Djurab, Chad, a 2.500 Km del Rift Valley. Asociada a la mandíbula, se halló un UP3 (KT12/H2). Los restos se han datado mediante correlaciones faunísticas en 3,4-3 Ma. El anñalisis inicial atribuyó los restos en Au. afarensis, pero varios rasgos derivados, sobre todo la forma de la sínfisis, más vertical, sirvieron para definir una nueva especie. La sínfisis mandibular es no presenta un torus marcado (parecido a Homo), y es parabólica. La parte anterior del mentón es casi vertical y escasamente reforzada. El cuerpo mandibular es grácil y la arcada dental ancha, corta y parabólica. Los dientes tienen el esmalte grueso, las coronas de incisivos y caninos son altas, con raíces largas. Los premolares tienen tres raíces. El P3 tiene dos cúspides y tres raíces, y el P4 está molarizado. Los UP3s son asimétricos y tienen tres raíces. Tener premolares con tres raíces lo diferencian de Au. afarensis, que tiene una o dos raíces en P3 y tres en P4.
- Brunet M, Beauvilain A, Coppens Y, Heintz E, Moutaye AH, Pilbeam D (1995) The first australopithecine 2,500 kilometres west of the Rift Valley (Chad) Nature 378(6554), 273-275.
- Guy F, Mackaye HT, Likius A, Vignaud P, Schmittbuhl M, Brunet M (2007) Symphyseal shape variation in extant and fossil hominoids, and the symphysis of Australopithecus bahrelghazali. J Hum Evol 55(1), 37-47.
- Lebatard AE, et al (2008) Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. Proc Natl Acad Sci USA 105(9), 3226-31.
- Lee-Thorp J, Likius A, Mackaye HT, Vignaud P, Sponheimer M, Brunet M (2012) Isotopic evidence for an early shift to C₄ resources by Pliocene hominins in Chad. Proc Natl Acad Sci USA 109(50), 20369-20372.
Kenyanthropus platyops
Lomekwi, Kenia (3,6-3,25 ma)
Esta especie se define a partir del cráneo KNM WT 40000 encontrado en 1998-1999 por el equipo de Meave G. Leakey en el miembro Kataboi, Lomekwi, formación Nachukui, datado en unos 3,5 ma, en los drenajes de los ríos Lomekwi y Topernawi, en la orilla oeste del lago

KNM-WT 40000 (a-d), KNM-WT 38350 (e, lateral), KNM-WT 40001 (f, lateral; g, inferior). Escala a-c 3 cm; d-g 1 cm.
Turkana, al norte de Kenia. El cráneo se diferencia de A. afarensis en la posición anterior de la raíz del arco zigomático, el plano transversal y sagital subnasal tienen una escasa proyección más allá de los caninos, el conducto auditivo externo es más pequeño y el segundo molar es más pequeño. KNM-WT40000 es un cráneo completo reconstruido a partir de cientos de piezas, alargado y con la cara plana y dientes pequeños. Capacidad craneal similar a la de los australopitecinos. Se ha sugerido que presenta ciertas similitudes con ER 1470, clasificado como Homo rudolfensis. La raíz anterior del zigoma está situada encima entre P3 y P4 como en la mayoría de los australopitecos. Es poco prognato con el clivus nasoalveolar, las paredes verticales de la abertura nasal y los molares en el mismo plano. El orificio nasal es alto y estrecho, en contraste con A. africanus y P. robustus. Carece de los pilares faciales anteriores presentes en estas dos especies. Es cráneo presenta una mezcla de caracteres primitivos (cráneo pequeño, hueso temporal, premolares y molares con esmalte de grosor similar a los del A. afarensis) y otros derivados (cara plana, dientes pequeños. Destaca también el fosil KNM-WT 38355, un fragmento de maxilar superior con dentición. Además se ha descrito un temporal, otra maxila, 3 fragmentos de mandíbula y 44 dientes aislados.
No está claro que K. platyops sea una especie diferenciada de A. afarensis, con la que se solapa geográficamente y cronológicamente.
Leakey MG, Spoor F, Brown FH, Gathogo PN, Kiarie C, Leakey LN, McDougall I (2001) New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433-440.
- Spoor F, Leakey MG, Leakey LN (2010) Hominin diversity in the Middle Pliocene of eastern Africa: the maxilla of KNM-WT 40000. Philos Trans R Soc Lond B Biol Sci 365(1556), 3377-3388.
Australopithecus garhi
Bouri, Etiopía (2,5 ma)
BOU-VP 12/130
El equipo de Tim White descubrió el cráneo BOU-VP 12/130 en el miembro Hatayae (Hata) dela península de Bouri (Middle Awash, Etiopía) entre 1990-1998. Los restos están datados en 2,5 ma. Garhi significa sorpresa en la lengua de los afar. La nueva especie presenta una serie de rasgos que la diferencian de A. afarensis por el gran tamaño de la dentición poscanina y la forma del primer premolar superior. En 1990 en Matabaietu y Gamedah se hallaron: un pequeño fragmento de parietal izquierdo (GAM-VP-1/2), un corpus mandíbular izquierdo sin dientes (GAM-VP-1/1) y un extremo distal de húmero izquierdo (MAT-VP-1 / 1). En 1996, se recuperó una mitad proximal de cúbito (BOU-VP-11/1), una porción proximal de fémur y elementos asociados del antebrazo de un infantil (BOU-VP-12/1A-G). Este esqueleto parcial incluye también diáfisis de fémur izquierdo y el húmero derecho, radio, cúbito y peroné, una falange proximal del pie y la base de la parte anterior de la mandíbula, que podrían corresponder al mismo individuo. La falange del pie es similar la de A. afarensis en tamaño, longitud y curvatura. En 1997 se encontró una mandíbula bastante completa con dientes (BOU-VP-17/1) y un cráneo parcial (frontal, parietal y maxilar con dientes) (BOU-VP-12/130) y un fragmento de bóveda craneal de otro individuo (BOU-VP-12/87). En 1998 se halló una diáfisis humeral (BOU-VP-35/1). El cráneo BOU-VP 12/130 recuerda a A. afarensis en la parte anterior del rostro (muy prognato, con superficie subnasal convexa). Su anatomía craneal y facial es primitiva, con grandes músculos faciales. El arco zigomático se inicia por encima de P4/M1 mientras que en otros australopitecinos lo hace en P3/P4. La vonstricción postoorbitaria es grande, con un cráneo redondeado com 450 cc de capacidad y cresta sagital. La arcada dental tiene forma de U, con brazos ligeramente divergentes, e los dientes son grandes. Se parece a Paranthropus aethiopicus en la parte distal de la arcada dental, pero los premolares no están completamente molarizados aunque su corona dibuja un perfil ovalado.
Australopithecus garhi se ha relacionado con el origen del género Homo en África del este, no por su similitud anatómica sino porque se ah encontrado asociado a huesos de bóvidos con marcas de descarnación realizadas con algún tipo de indústria lítica, así como un fémur de caballo encontrado en el mismo nivel. Estos huesos proporcionan evidencias claras de el uso de indústrias líticas hace 2,5 ma en Bouri using, y el único hominino encontrado en este yacimiento es Australopithecus garhi. Sin embargo, existen notables similitudes de este especimen con A. africanus de sudáfrica.
- Asfaw B, White T, Lovejoy O, Latimer B, Simpson S, Suwa G (1999) Australopithecus garhi: a new species of early hominid from Ethiopia. Science 284(5414), 629-35. Erratum in: Science 1999 284(5420), 1623. Science 2004 304(5671), 684.
- Heinzelin J de, Clark JD, White T, Hart W, Renné P, WoldeGabriel G, Beyene Y, Vrba E (1999) Environment and behavior of 2.5-million year old Bouri hominids. Science 284, 625-629.
Australopithecus africanus

Australopithecus africanus
Esta especie sólo se ha descrito en Sudáfrica en los yacimientos de Sterkfontein, Makapansgat y Taung. Algunos autores destacan similitudes entre A. africanus y A. garhi de África del este y tambié se sugerido la existencia de A. africanus dental (similitudes morfológicas de los dientes en el yacimiento de Omo en Etiòpia (National Geographic, Febrero 1997). La anatomía craneofacial de A. africanus bastante derivada (frente algo verticalizada, calota craneal relativamente corta y alta, prognatismo facial reducido) ha sugerido que podría estar en la base de la emergència de Homo, aunque sus rasgos son claramente australopitecinos. Tendría extremidades anteriores proporcionalmente más largas que en humanos actuales (índice húmero-femoral 76,9-96,8). La curvatura de su columna es parecida a la de los humanos moderns, aunque los cuerpos vertebrales lumbares eran pequeños. Sería un bípedo facultativo, de aproximadamente 1,2 m de estature, Sts-14) y 53 Kg de peso. La cronologia de A. africanus es poco precisa, entre 3,5 y 2,3 ma. El holoyipo es Taung 1 (el Niño de Taung) descrito por Raimon Dart en 1924, datado en 2,3 ma, con una edad de unos 3 años y una capacidad craneal de 405 cc (440 cc adult). En 1936 se descubrieron nuevos fósiles en Skertfontein (TM 1511 y TM 1512) y en 1947 los de STS 5. Además se han encontrado restos en Makapansgat y Gladysvale.
Makapansgat,Sudáfrica (3-2,6 ma)
Los materiales fósiles encontrados en Limeworks Cave, en los miembros 3 y 4, Northern Province (Sudáfrica) están datados en 3-2,6 ma. Desde 1946 se han encontrado 27 especímenes de al menos 10 individuos en un entorno con un mosaico de hábitats. El estudio de isótopos estables refleja un consumo de hierbas y juncos (C4) (Sponheimer y Lee-Thorp, 1999). Entre los fósiles más significativos astán MLD 1 (calota parcial de adulto), MLD 2 (mandíbula), MLD 6/23 (mandíbula), MLD 9 (mandíbula), MLD 11/30 (maxilar y dientes), MLD 18 (mandíbula), MLD 37/38 (fragmento de cráneo), MLD 40 (mandíbula).
Taung, Sudáfrica (3-2,6 ma)
El niño de Taung (Taun 1)
El niño de Taung (Taung 1) se descubrió en 1924 (Raymond Dart). El fósil incluye parte del cráneo (molde endocraneal), la cara y la mandíbula de un individuo joven (3-4 años), con la dentición decidua (Kuhn et al. 2016, Palaeontologia africana 51, 10–26) y el primer molar permanente que empieza a emerger. Su capacidad craneana es de 410 cc y se supone que de adulto tendría una capacidad de 440 cc. El gran tamaño del cerebro y pequeño tamaño de los caninos, junto con la posición centrada del foramen mágnum sugirieron a Dart que era bípedo. Presenta otros rasgos llamativos: abertura nasal pequeña, frente verticalizada y torus supraorbital poco pronunciado.
Sterkfontein, Sudáfrica (2,6-2 ma)
Sterkfontein (término en afrikáans para manantial fuerte) es un conjunto de cuevas de piedra caliza en las que se han encontrado, desde finales del siglo XIX, numerosos de restos fósiles de homininos. Se localiza en la provincia de Gauteng, al noroeste de Johannesburgo. Los sitios arqueológicos de Swartkrans, Kromdraai y Wonder Cave se encuentran en esta misma área. Las excavaciones en las cuevas comenzaron a fines de la década de 1890, cuando mineros de piedra caliza notaron que había fósiles, lo que llamó la atención de los científicos. En 1936 empezaron las excavaciones planificadas por estudiantes del profesor Raymond Dart y el Dr. Robert Broom de la Universidad de Witwatersrand. En 1947 se encontró un cráneo casi completo de una hembra adulta de Australopithecus africanus. Broom le dio el nombre de Plesianthropus transvaalensis y lo apodó Mrs. Ples (2,6-2,8 ma). En 1997 Ronald J. Clark encontró un esqueleto casi completo el que denominó Little Foot, ya que las primeras partes encontradas en 1995 eran huesos de un pie de unos 3,3 ma. Las excavaciones continúan en la actualidad.

Sts 5 Australopithecus africanusSTS 1. Parietal
- STS 2. Maxilar y dientes asociados. Edad al morir 2,52 años (Smith et al, 2015).
- STS 5. La Señora Ples (Mrs. Ples). 1947. 2 ma. Robert Broom. Cráneo femenino muy bien conservado (Villmoare et al, 2013) de 485 cc.
- STS 14. 1947. 2,5 ma . Robert Broom y JT Robinson. Casi completa columna vertebral con 6 vértebras lumbares y otras torácicas, pelvis, fragmentos de costillas y un fragmento de fémur. Sts 5 y Sts 14 podrían corresponder al mismo individuo. Probable individuo masculino con estatura de 122-137 cm (Robinson, 1972).
- STS 17.
- STS 24. Maxilar y dientes mandibulares. Edad al morir 4,35 años (Smith et al, 2015).
- STS 71. Cráneo.
- STW 13.
- STW 183. Maxilar.
- STW 252. Cráneo.
- STW 352. Calcáneo (Zeininger et al, 2016).
- STW 384. Mandíbula.
- STW 431. Esqueleto parcial. Incluye seis vértebras lumbares (Odes et al, 2017).
- STW 432. Esqueleto parcial de tamaño grande.
- STW 498. Maxilar y mandíbula.
- STW 505. Cráneo. 500 cc.
- STW 573. Little Foot. 1994-1997. Gruta Silberberg. 2,6-2,2 ma. Ron Clarke. Esqueleto casi completo (Clarke, 2008 South African Journal of Science
104, 443-449. November/December 2008). El pie izquierdo conserva el astrágalo, escafoides, primer cuneiforme y mitad proximal del dedo gordo. La mano presente una articulación trapecio / primer metacarpiano diferente a la de humanos y chimpancés, un pulgar humano robusto y falanges proximales tan curvadas como las de Au. afarensis.
-
STW 670. Fragmento proximal de primera costilla derecha (Tawane et al, 2016).
- TM 1511. 1936.
- TM 1512. 1936. Robert Broom Se trata de partes de la cara, mandíbula inferior y bóveda del cráneo.
Gladysvale, Sudáfrica (2,5-1,7 ma)
Gladysvale está situado 13 km al nordeste de Sterkfontein, datados entre 2,4-2 ma, cerca de Johannesburgo en la región sur del Transvaal. Se han encontrado diversos dientes. Además se han encontrado numerosos restos de fauna Plio-Pleistocénica que indica condiciones ecológicas de savanaen un periodo datado entre 1,7 y 2,5 ma.
- Dart RA, Salmons A (1925) Australopithecus africanus: The Man-Ape of South Africa.
- Dart RA (1948) The Makapansgat proto‐human australo‐pithecus prometheus. Am J Phys Anthropol 6(3), 259-284.
- Dart RA (1949) Innominate fragments of Australopithecus prometheus. Am J Physi Anthropol 7(3), 301-334.
- Broom R (1949) Another new type of fossil ape-man» Nature 163, 57.
- Broom R, Robinson JT. Further evidence of the structure of the Sterkfontein ape-man Plesianthropus. Transvaal Museum, Pretoria. Part I(1950).
- Tobias PV (1974) Recent studies on Sterkfontein and Makapansgat and their bearing on hominid phylogeny in Africa. Goodwin Series 2, 5-11.
Berger LR, Keyser AW, Tobias PV (1993) Brief communication: Gladysvale: first early hominid site discovered in South Africa since 1948. Am J Phys Anthropol. 1993 Sep;92(1):107-11.
- Clarke RJ, Tobias PV (1995) Sterkfontein Member 2 foot bones of the oldest South African hominid. Science 269(5223), 521-524.
- Sponheimer M, et al (2005) Hominins, sedges, and termites: new carbon isotope data from the Sterkfontein valley and Kruger National Park. J Hum Evol 48(3), 301-312.
- Clarke R (2013) Australopithecus from Sterkfontein Caves, South Africa. The paleobiology of Australopithecus. Springer Netherlands, pp. 105-123.
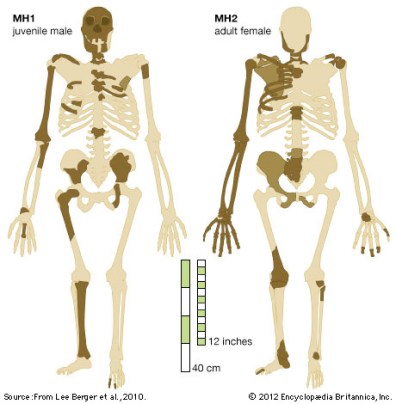
Australopithecus sediba
Malapa, Sudáfrica (1,977 ma)

MH1 – Au. sediba
Los restos atribuidos a esta especie fueron hallados por el hijo de Lee R. Berger, de la universidad de Witwatersrand en Johannesburgo (Sudáfrica) el 15 de agosto de 2008 en Malapa, a 15 Km de Sterkfontein, Swartkrans, y Kromdraai. Los restos de Australopithecus sediba (MH1 y MH2) aparecieron en un pozo con sedimentos resultado de la erosión de un sistema antiguo de cuevas profundas, en la Facies D, datada en 1,977 ± 0.002 ma. La diversidad de fauna, la abundancia de especímenes completos y/o articulados y la falta de huesos modificados por carnívoros indican que los animales quedaron atrapados en el sistema de cuevas. Las mayoría de fracturas visibles en MH1 y MH2 son post-deposicionales, pero las del miembro superior derecho de la hembra adulta junto con posibles fracturas en la mano, indican claramente la resistencia activa a un impacto, mientras que las de la mandíbula del juvenil son consistentes con un golpe en la cara. La presencia de estos traumatismos apoya la hipótesis de la caída y muerte de un australopitecino por causas diferentes a la depredación o la muerte natural.
Los restos de de los hominini encontrados pertenecen a una hembra adulta y a un joven, sin poder encuadrarlos en taxones existentes. Por ello, definieron una nueva especie, Australopithecus sediba. Sediba significa manantial inagotable en la lengua seSotho. Las diferencias entre ambos especímenes indican un amplio rango de variación morfolófica de la especie. Comparte con Australopithecus africanus numerosas características en la bóveda del cráneo, el esqueleto facial, la mandíbula y los dientes. Pero otras características (dientes más pequeños, pómulos menos pronunciados, nariz prominente, piernas más largas, rasgos de la pelvis) lo acercan a Homo. No obstante, en base a algunos rasgos entre los que destaca el pequeño tamaño del cerebro, se ha incluido dentro del género Australopithecus. El holotipo de la especie es MH1, un joven (12-13 años) representado por un cráneo parcial, un fragmento de mandíbula y parte del esqueleto postcraneal. Su volumen cerebral (420 cc) se estima alcanzaba el 95% del tamaño adulto. El paratipo es MH2, una hembra adulta representada por un diente maxilar, una mandíbula parcial y parte del esqueleto postcraneal. Del análisis de las manos, con dedos cortos y pulgar relativamente largo, capaces de realizar la pinza de precisión, puede inferirse la capacidad para producir herramientas líticas.Los largos brazos y la estructura de los pies sugieren capacidad trepadora. Se deduce que esta especie tenía una amplia variedad de comportamientos y podía explotar diversos entornos. Se estima que la altura máxima de la especie era 1,30 m. En MH1 y MH2 se han encontrado restos de tejidos blandos.
Australopithecus sediba
Diversas hipótesis filogenéticas son posibles: 1) Australopithecus sediba probablemente desciende de A. africanus, y parece el australopiteco más cercano a Homo. Tiene rasgos craneodentales y patrones de locomoción más avanzados que Sterkfontein Stw 53, Olduvai OH 62 y Koobi-Fora KNM-ER 3735, más o menos contemporáneos en el tiempo y asignados a H. habilis, por lo que esta asignación se vuelve incierta. 2) Su talón, más primitivo que el de Australopithecus afarensis, sugiere que forma parte de un linaje diferente al representado por A. afarensis y A. africanus, pero el patrón de crecimiento facial es único entre los Hominini tempranos. 3) Australopithecus sediba podría estar en el camino evolutivo a Homo ergaster. Esta alternativa es defendida, entre otros, por Chris Stringer. Homo habilis constituiría una rama aislada y sin descendencia del árbol de Homo. 4) Si AL-666-1 no es Homo y Stw 53, OH 62 y KNM-ER 3735 no son H. habilis, entonces A. sediba sería anterior y se podría admitir una secuencia filogenética Australopithecus sediba – Homo habilis – Homo erectus. Sediba comparte con H. erectus algunos rasgos craneales que presenta H. habilis. Contradice esta hipótesis que la anatomía de los restos candidatos a Homo temprano es diferente a la del sediba. El fósil de Homo temprano LD 350-1 es más derivado en su morfología dental y mandibular a pesar de ser anterior en el tiempo en 0,75-0,8 ma. 5) Podría representar un grupo hermano que persistió durante algún tiempo después de la primera aparición de Homo. Ésta parece la hipótesis más probable, defendida entre otros por Donald Johanson. En todo caso, A. sediba prueba que la transición entre Australopithecus y Homo se produjo en forma de mosaico, con cambios en la pelvis, aumento de la distancia de las líneas temporales y reducción de la constricción postorbital antes de que aumentara la encefalización. La reducción dental fue anterior a la reorganización de cúspides.
- Berger LR, de Ruiter DJ, Churchill SE, Schmid P, Carlson KJ, Dirks PH, Kibii JM (2010) Australopithecus sediba: a new species of Homo-like australopith from South Africa. Science 328(5975), 195-204.
- Dirks Paul HGM, et al (2010) Geological setting and age of Australopithecus sediba from southern Africa. Science 328(5975), 205-208.
- Pickering R, et al (2011) Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo. Science 333(6048), 1421-1423.
- Henry AG, et al (2012) The diet of Australopithecus sediba. Nature 487(7405), 90-93.
Prang TC (2015) Rearfoot posture of Australopithecus sediba and the evolution of the hominin longitudinal arch. Sci Rep 5, 17677.
Lacruz RS, Bromage TG, O’Higgins P, Toro-Ibacache V, Warshaw J, Berger LR (2015) Distinct growth of the nasomaxillary complex in Au. sediba. Sci Rep 15, 15175.
Ritzman TB, Terhune CE, Gunz P, Robinson CA (2016) Mandibular ramus shape of Australopithecus sediba suggests a single variable species.J Hum Evol 100, 54-64.
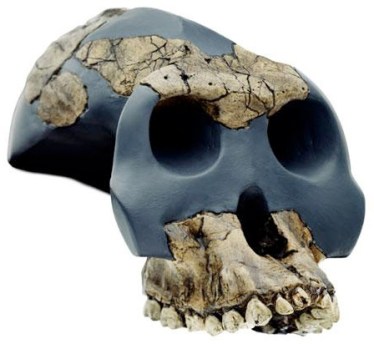
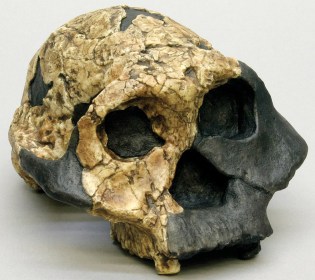
Paranthropus aethiopicus

WT 17000 – Paranthropus aethiopicus
En 1967 Camille Arambourg e Yves Coppens encontraron en uno de los yacimientos del curso bajo del río Omo, cerca del lago Turkana, en Etiopía, una mandíbula (Omo 18) de unos 2,6 millones de años de antigüedad, y en 1985 el paleoantropólogo estadounidense Alan Walker descubrió un cráneo (el cráneo negro) perteneciente a la misma especie. Este fósil permitió definir esta nueva especie. Algunas características de este cráneo recuerdan al de Australopithecus afarensis: base del cráneo plana, articulación de la mandíbula baja, marcado prognatismo facial y cerebro pequeño. Se ha propuesta que P. aethiopicus sería en ancestro común, datado en 2,5 ma, de Paranthropus boisei en África del este y Paranthropus robustus en Sudáfrica, que muestran una base del cráneo inclinada, una articulación de la mandíbula profunda, una cara plana y un cerebro algo mayor (de 500 a 545 cc). Estas características son compartidas con los primeros representantes del género Homo y se han atribuido a evolución paralela (homoplasia). Actualmente se acepta que el linaje robusto del Este de África evolucionó localmente: A. afarensis ⇒ P. aethiopicus ⇒ P. boisei. En cambio, se ha sugerido que las similitudes dentales y del aparato masticador de P. robustus y P. boisei se deben a evolución paralela.
Formación Shungura, Río Omo, Etiopía (2,3-2,6 ma)
El holotipo de esta especies es el especimen Omo 18-1967-18, un fragmento de mandíbula datado en 2,7 ma, hallado por Camille Arambourg y Yves Coppens en el Miembro C de la Formación Shungura, en Etiopía. Otros restos significativos de este yacimiento (hallados entre 1902-1976) son: las mandíbulas Omo 44-1970-2466, Omo 57-4-1968-41, Omo 860-2 y Omo L55-s-33, el cráneo Omo L338-y-6 y 20 dientes dientes postcaninos inferiores, algunos pertenecientes a las mandíbulas señaladas.
Formación Nachukui, West Turkana, Kenya (2,36-2,57 ma)
En la Formación Nachakui, cuenca del Lomekwi, al Oeste del lago Turkana, Alan Walker y sus colaboradores del equipo de Richard Leakey encontraron en 1985 el fósil más completo de esta especie, KNM-WT 17000 (Black Skull), un cráneo sin dientes de 2,5 ma, (es el único cráneo conocido de esta especie), cuyo color oscuro se debe a la presencia de minerales de manganeso. Presenta un aparato masticatorio masivo, con un esqueleto facial muy ancho y gran prognatismo facial, arcos zigomáticos muy abiertos, superficie del cigomático inclinada hacia delante, cráneo bajo, con capacidad craneal baja (410 cc.), líneas temporales marcadas y cresta sagital robusta. También destaca el especimen KNM-WT 16005, un fragmento de mandíbula encontrado también en 1985.
Ndolanya Beds, Laetoli, Tanzania (2,5-2,7 ma)
Un maxilar superior sin deintes: EP 1500/01.
- Arambourg C, Coppens Y (1968) Decouverte d’un Australopithecien nouveau dans les gisements de l’Omo (Ethiopia). South African Journal of Science 64, 58-59.
- Olson TR (1985) Cranial morphology and systematics of the Hadar formation hominids and Australopithecus africanus. En: Delson E, Ancestors: the Hard Evidence. Nueva York: Alan R Liss: 102–119.
- Walker A, Leakey REF, Harris JM, Brown FH (1986) 2.5-Myr Australopithecus boisei from west of Lake Turkana, Kenya. Nature 322, 491-493.
Paranthropus boisei
La especie Paranthropus boisei fue definida por Louis Leakey en 1959. Lo fósiles de esta especie, datados entre 2,3 y 1,2 ma, proceden de África del Este. Los materiales anteriores a 2,3 ma correspondientes a homínidos robustos se clasifican dentro del taxóm P. aethiopicus. El primer Paranthropus boisei apareció en el yacimiento BK, Lower Bed II, de la Garganta de Olduvai, en 1955: OH3 (Olduvai Hominid 3), con caninos y un molar decidua. En 1959 Mary y Louis Leakey descubrieron OH5, datado en 1,8 ma, conocido como Dear boy (Cascanueces). Se trata de un cráneo de subadulto bien preservado, casi completo, de 530 cc. En la garganta de Olduvai no se halló ninguna mandíbula, pero en 1964 apareció en la orilla oeste del Lago Natron una mandíbula adulta bien preservada, robusta, con megadoncia postcanina (NMT-W64-160, Peninj 1). En 2013, Manuel Domínguez-Rodrigo publicó el hallazgo en 2010 en BK de un conjunto catalogado como OH 80, datado en 1,338 ± 0.024 ma, que es el primer esqueleto parcial hallado en África del Este, clasificado en el taxón Paranthropus boisei de forma segura. El especimen KNM-ER 1500 (fragmentos postcraneales de Koobi Fora, Kenya) fue asignado a P. boisei por su asociación espacial con un fragmento de mandíbula. Otros restos relacionado con esta especie incluyen: KNM-ER 406. 1969. Turkana, Kenia. 1,7 ma. Richard Leakey. Se trata de un cráneo completo al que sólo le faltan los dientes. Su capacidad craneal es de 510 cc; KNM-ER 732. 1970. Turkana. 1,7 ma. Richard Leakey. El cráneo es similar al OH 5, pero más pequeño con cresta sagital. Capacidad craneana de 500 cc; KGA10-525. 1993. Konso Etiopia. 1,4 ma. A. Amzaye. Se trata de un cráneo, incluyendo una mandíbula. Su capacidad craneana es de 545 cc.
Paranthropus boisei se caracteriza por su grandes robustez craneofacial y por su megadoncia. Es más robusto que los parántropos de Sudáfrica. Las coronas de los premolares mandibulares presentan un talónido particularmente largo y elaborado (Suwa, 1988); la corona del primer molar mandibular tiene cúspides adicionales confinadas al talónido (Wood y Abbott, 1983). Es el único hominino que combina una cara amplia, masiva, plana, con megadontia postcanina y un modesto volumen endocraneal (400-545 cc) (Bilsborough y Wood, 1988). El cerebro del P. boisei no creció a lo largo de los más de 1 ma que existió la especie (J. Hawks, 2011). La cresta sagital es enorme, los parietales son relativamente largos y estrechos, el occipital bajo y amplio y los huesos de la bóveda craneal son muy finos. Presenta una reducción del tamaño de caninos e incisivos, en línea recta delante del paladar. El tamaño del tercer molar superior es más pequeño que el segundo molar. La especies muestra un gran dimorfismo sexual en tamaño y robustez (las hembras no presentan cresta sagital).
Los restos postcraneales de OH 80 sugieren que P. boisei habría tenido una estatura de 156 ± 3,91 cm y un peso entre 40-61,7 Kg. Esto supone que un indiviuo sustancialmente más grande que el presuntamente femenino KNM-ER 1500, lo que indica un alto grado de dimorfismo sexual. Las dimensiones de OH 80-12 quedan dentro del rango de variación de H. erectus, pero muestran algunas características que reflejan las diferencias en la biomecánica de las extremidades inferiores de P. boisei y H. erectus.Los restos de OH 80-11 sugieren que P. boisei tenía las extremidades superiore relativamente largas y poderosas; es el hueso del antebrazo más robusto conocido del Plioceno y comienzos del Pleistoceno. Es posible que, perdidas las adaptaciones para la braquiación con los primeros Paranthropus, en P. boisei surgiesen de nuevo como una apomorfia. P. boisei sería una especie altamente especializada craneodentalmente, simpátrica con Homo, con un esqueleto postcraneal robusto y con bipedación terrestre combinada con arboricolismo ocasional.
West Turkana, Kenia (2,3-1,6 ma)
KNM-WT 17396. 1.65–1.79 ma. M3 (o M2) mandibular izquierdo. KNM-WT 17400. 1.67–1.87 ma. Craneo anterior y endocasto. KNM-WT 18600. 1.65–1.79 ma. P3 maxilar izquierdo. KNM-WT 37100. 1.65–1.79 ma. M3 mandibular izquierdo. KNM-WT 37744. 1.65–1.79 ma. Canino maxilar izquierdo. KNM-WT 37747. 1.65–1.79 ma. M1 maxilar izquierdo. KNM-WT 37748. 1.65–1.79 ma. M3 maxilar derecho
Koobi Fora, Kenia (2,2-1,4 ma)

ER 406
- KNM-ER 403. 1.67 ma. Fragmento mandibular derecho.
- KNM-ER 404. 1.49–1.55 ma. Fragmento mandibular derecho.
- KNM-ER 405. 1.87 ma. Fragmentos palatomaxilares.
- KNM-ER 406. 1.56–1.60 ma. Cráneo sin dientes.
- KNM-ER 407. 1.85 ma. Calvaria.
- KNM-ER 725. 1.49–1.55 ma. Cuerpo mandibular izquierdo.
- KNM-ER 726. 1.55–1.65 ma. Cuerpo mandibular izquierdo.
- KNM-ER 727. 1.56–1.60 ma. Fragmento mandibular derecho.
- KNM-ER 728. 1.49–1.55 ma. Cuerpo mandibular derecho.
- KNM-ER 729. 1.49–1.55 ma. Mandíbula.
- KNM-ER 732. 1.56–1.60 ma. Cráneo parcial.KNM-ER 733. 1.49–1.55 ma. Fragmentos de cráneo. KNM-ER 801. 1.56–1.60 ma. Fragmento de mandíbula.
- KNM-ER 802. 1.56–1.60 ma. Isolated teeth.
- KNM-ER 805. 1.49–1.55 ma. Fragmento de mandíbula
ER 732
- KNM-ER 810. 1.77 ma. Fragmento de mandíbula.
- KNM-ER 812. 1.77 ma. Fragmento de mandíbula.
- KNM-ER 814. 1.77 ma. Fragmentos de bóveda craneal.
- KNM-ER 816. 1.77 ma. Canino maxilar.
- KNM-ER 818. 1.55–1.65 ma. Fragmento de mandíbula.
- KNM-ER 1170. 1.56–1.60 ma. Fragmentos de bóveda craneal.
- KNM-ER 1171. 1.56–1.60 ma. Diente aislado.
- KNM-ER 1467. 1.49–1.55 ma. Diente aislado.
- KNM-ER 1468. 1.55–1.65 ma. Fragmento de mandíbula.
- KNM-ER 1469. 1.87–1.90 ma. Fragmento de mandíbula.
- KNM-ER 1477. 1.85–1.90 ma. Mandíbula sin ramis.
- KNM-ER 1479. 1.87 ma. Diente aislado.
- KNM-ER 1500. 1.87–1.90 ma. Esqueleto parcial.
- KNM-ER 1509. 1.56–1.60 ma. Diente aislado.
- KNM-ER 1803. 1.87–1.90 ma. Fragmento de mandíbula. KNM-ER 1804. 1.77 ma. Fragmento de mandíbula.
- KNM-ER 1806. 1.85 ma. Fragmentos de mandíbula.
- KNM-ER 1816. 1.56–1.60 ma. Fragmentos de mandíbula.
- KNM-ER 1818. 1.56–1.60 ma. Diente aislado.
- KNM-ER 1819. 1.60–1.87 ma. Diente aislado.
- KNM-ER 1820. 1.67–1.70 ma. Fragmentos de mandíbula.
- KNM-ER 2607. 1.60–1.87 ma. Diente aislado.
- KNM-ER 3229. 1.85 ma. Fragmento de mandíbula.
- KNM-ER 3230. 1.56–1.60 ma. Mandíbula sin ramas.
- KNM-ER 3729. 1.87–1.90 ma. Fragmento de mandíbula.
- KNM-ER 3731. 1.87–1.90 ma. Fragmentos de mandíbula.
- KNM-ER 3737. 1.56–1.60 ma. Diente aislado.
- KNM-ER 3885. 1.77–1.79 ma. Diente aislado.
- KNM-ER 3886. 1.77 ma. Diente aislado.
- KNM-ER 3887. 1.50–1.65 ma. Diente aislado.
- KNM-ER 3889. 1.49 ma. Fragmento de mandíbula.
- KNM-ER 3890. 1.56–1.60 ma. Diente aislado.
- KNM-ER 3952. 1.87 ma. Diente aislado.
- KNM-ER 3954. 1.60–1.87 ma. Fragmentos de mandíbula.
- KNM-ER 5429. 1.56–1.60 ma. Fragmento de mandíbula.
- KNM-ER 5679. 1.56–1.60 ma. Diente aislado.
- KNM-ER 5877. 1.49–1.55 ma. Fragmento de mandíbula.
- KNM-ER 6080. 1.55–1.65 ma. Diente aislado.
- KNM-ER 6082. 1.55–1.65 ma. Diente aislado.
- KNM-ER 6128. 1.77 ma. Diente aislado.
- KNM-ER 13750. 1.87 ma. Cráneo con endocasto.
- KNM-ER 15930. 1.77 ma. Mandíbula.
- KNM-ER 15940. 1.77 ma. M2 o M3 germinales izquierdos y derechos.
- KNM-ER 15950. 1.49–1.55 ma. M3 mandibular muy erosionado.
- KNM-ER 16841. 1.60–1.87 ma. Mandíbula sin dientes.
- KNM-ER 17760. 1.77 ma. I2 maxilar izquierdo.
- KNM-ER 23000. 1.87 ma. Cráneo.
- KNM-ER 25520. 1.80–1.90 ma. Cuerpo mandibular derecho.
Chesowanja, Kenia (1,5 ma)
- KNM-CH 1. 1.42 ma. Mitad del rostro y base craneal anterior.
- KNM-CH 302. 1.42 ma. Fragmentos de molar.
- KNM-CH 304. 1.42 ma. 5 fragmentos craneales
Chiwondo Beds, Malawi (2,5-2,3 ma)
- RC 911. 2.3–2.5 ma. Fragmento de maxilar
Omo, Etiopía (2,3-1,2 ma)
Omo L7A-125
- Omo F203-1. 1.40–1.45 ma. M3 mandibular derecho.
- Omo L7A-125. 2.19–2.27 ma. Mandíbula.
- Omo L40-19. 2.36–2.40 ma. Cúbito derecho.
- Omo L74A-21. 2.19–2.27 ma. Mandíbula derecha.
- Omo L427-7. 1.8–2.0 ma. Mandíbula derecha.
- Omo L628-1. 2.27–2.33 ma. P4 maxilar izquierdo.
- Omo L628-2. 2.27–2.33 ma. M3 mandibular derecho.
- Omo L628-3. 2.27–2.33 ma. M3 mandibular izquierdo.
- Omo L628-4. 2.27–2.33 ma. P4 mandibular izquierdo.
- Omo L628-5. 2.27–2.33 ma. P4 mandibular derecho.
- Omo L628-9. 2.27–2.33 ma. M1 mandibular izquierdo.
- Omo L704-2. 2.40–2.52 ma. P3 deciduo mandibular izquierdo.
- Omo L726-11. 2.19–2.27 ma. P4 maxilar derecho.
- Omo L797-1. 2.19–2.27 ma. Fragmento de P4 mandibular izquierdo.
- Omo 47-46. 2.19–2.27 ma. M2 desgastado mandibular derecho.
- Omo 47-1500. 2.10 ma. M2 mandibular derecho.
- Omo 76-37. 2.27–2.33 ma. Mitad bucal de M3 mandibular izquierdo.
- Omo 141-2. 2.27–2.33 ma. M1 o M2 Maxilar M1 or M2.
- Omo 323-896. 2.19–2.27 ma. Cráneo parcial
Konso, Etiopía. 1,4 ma.
En 1997 el equipo de Gen Suwa halló un cráneo asociado a una mandíbula.
- KGA 10-506. 1.41–1.43 ma. Paladar izquierdo con dentición.
- KGA 10-525. 1.41–1.43 ma. Cráneo parcial.
- KGA 10-565. 1.41–1.43 ma. M1 maxilar derecho.
- KGA 10-570. 1.41–1.43 ma. Mandíbula juvenil.
- KGA 10-900. 1.41–1.43 ma. Fragmentos molares.
- KGA 10-1455. 1.41–1.43 ma. Parietal izquierdo.
- KGA 10-1720. 1.41–1.43 ma. M3 mandibular izquierdo.
- KGA 10-2705. 1.41–1.43 ma. M2 mandibular derecho.
- KGA 10-2741. 1.41–1.43 ma. Fragmentos molares
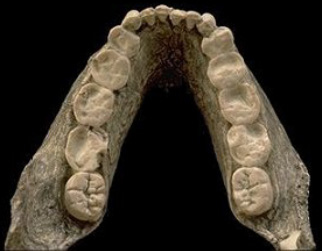
Olduvai, Tanzania (1,8-1,2 ma)

OH 5
- OH 3. 1.40–1.45 ma. Molar aislado.
- OH 5. 1.79–1.85 ma. Cráneo.
- OH 20. 1.66–1.79 ma. Fragmento proximal del fémur izquierdo.
- OH 26. 1.66–1.79 ma. Molar maxilar sin erupcionar.
- OH 30. 1.66–1.79 ma. Dientes deciduos y permanentes.
- OH 36. 1.40–1.45 ma. Cúbito casi completo.
- OH 38. 1.40–1.45 ma. M2 mandibular and dos incisivos derechos.
- OH 46. 1.79–1.85 ma. Fragmento de corona premolar o molar.
- OH 80-1. I2 inferior derecho casi competo.
- OH 80-2. Fragmento de P3 superior derecho unido a un pequeño fragmento de maxilar.
- OH 80-3. Fragmento de P4 superior izquierdo.
- OH 80-4. Fragmento de M1 superior izquierdo.
- OH 80-5. Fragmento de maxilar izquierdo de 5 cm mesiodistalamente, que conserva I1 e I2.
- OH 80-6. I1 superior derecho. OH 80-7. I2 superior derecho. OH 80-8 y OH 80-9. Raíces extremadamente robustas sin coronas.
- OH 80-10. Porción distal de húmero izquierdo.
- OH 80-11- Fragmento proximal de radio derecho con la mayor parte de su eje.
- OH 80-12. Segmento de la diáfisis de un fémur derecho.
- OH 80-13. Fragmento de eje de una tibia.
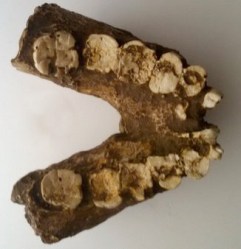
NMT-W64-160 – Peninj
Peninj, Tanzania (1,7 ma)
- NMT-W64-160. 1.56–1.70 ma. Mandíbula
- Leakey LSB (1959) A new fossil skull from Olduvai. Nature 184(4685), 491-493.
- Fleischer RL et al (1965) Fission-Track Dating of Bed I, Olduvai Gorge. Science 148(3666), 72-74.
- Leakey MD (1967) Olduvai gorge. Eds. Phillip V. Tobias, and L. S. B. Leakey. Vol. 2. Cambridge University Press.
- Blumenschine RJ, Stanistreet IG, Masao FT (2012) Olduvai Gorge and the Olduvai Landscape Paleoanthropology Project. J Hum Evol 63(2), 247-250.
Paranthropus robustus
Esta especie, datada entre 2,2–1 ma, fue definida por Robert Broom en 1938 en base al descubrimiento del especimen TM 1517 en el yacimiento de Kromdraai, el único esqueleto parcial de esta especie conocido hasta ahora (Broom, 1938a, Broom 1938b, Broom 1942 y Broom, 1943). Estatura 1,32-1,10 m, algo superior a la de Australopithecus. Peso 40-32 kg. Bipedación casi humana. Los restos de las manos indican mayor precisión que la de los australopitecinos. Paranthropus robustus muestra un alto dimorfismo sexual.
Rasgos anatómicos del aparato masticador:
- Grandes y anchos molares y premolares. Caninos e incisivos pequeños. Mandibulas masivas, cortas, altas y extremadamente robustas con forma de U divergente (mastiación potente). No tanto como P. boisei. Corpus mandibular amplio y profundo con una rama mandibular alta, relativamente vertical. No se descarta que, aparte de la dieta vegetal, ingiriese insectos. Según los estudios de Andrew Sillen y Julia Lee-Thorp, P. robustus ingería proteínas animales.
- Caja craneal bastante alta y desarrollada, con cresta sagital. Volumen cerebral 530 cc. Enorme hueso malar. Rostro recto, con arco zigomático muy ancho y robusto. Torus supraorbitario masivo.
- Esmalte dental muy grueso, mucho más que el A. africanus. Según Wynn, Cerling y Sponheimer (2013) su dieta estaba compuesta de alimentos procedentes de árboles y arbustos, pero también de hierbas y juncos, con gran variación entre individuos. Según Ungar y Hlusko (2016), a pesar de su aparatosa dentadura, solo consumía alimentos duros ocasionalmente.
- Los australopitecinos robustos del Este de África también tienen esmalte muy grueso en los molares. Podría ser una sinapomorfia, lo que implicaría una rama robusta común antes de la separación. Pero también podría ser un rasgo desarrollado por separado en cada especie (homoplasia).
Kromdraai B (KB), Gauteng, Sudáfrica (2-1,5 ma)
Situado a 2 Km al este de Sterkfontein. Se ha sugerido que algunos especímenes de Kromdraai datan del origen del género Paranthropus (Kaszycka 2002; Tobias 1988) y son intermedios entre los hominini de Makapansgat Miembros 3/4 y Sterkfontein Miembro 4, pero más derivados que Swartkrans. Las excavaciones fueron dirigidas por Broom (1938-1944), Brain (1955-1956), Vrba (1977-1980) y Thackeray (1993-2002). Desde 2002, las excavaciones están a cargo del Kromdraai Research Project. Los contextos para las excavaciones de Broom y Brain son confusos, imposibilitando una datación precisa. Los carnívoros fueron el principal agente de acumulación.
- Individuo 1. Subadulto. TM 1517. Broom (1938a); Broom y Schepers (1946); Broom (1943); Broom (1938b); Day y Thornton (1986); Day (1978); Skinner et al. (2013); Broom (1942).
- Individuo 2. Juvenil, 1,63-2,02 años (Smith et al, 2015). TM 1536. Fragmento mandibular izquierdo con I1, I2, di2, dm1-2, M1; L dC. Broom (1941); Broom y Schepers (1946).
- Individuo 3. Adulto. TM 1600. Dos fragmentos del corpus mandibular, con M2-M3 (fragmento1) y P3 (fragmento 2). Brain (1981).
- Individuo 4. Juvenil. TM 1601. dm1 inferior derecha, P3, C, P4 germinales inferiores derechos, M1 germinal superior izquierdo, dc inferior derecho. Brain (1981).
- Individuo 5. Adulto. TM 1602. Fragmento maxilar derecho con raíces de P4 a M3. Brain (1981).
- Individuo 6. Adolescente. TM 1603. M3 superior izquierdo. Dump, 1944; Broom and Schepers, 1946.
- Individuo 7. TM 1604. Juvenil. dm2 inferior izquierdo. Brain (1981).
- Individuo 8. Adulto. TM 1605. Cadera izquierda. Brain (1981); Robinson (1972).
- Individuo 9. Adulto. KB 5063. M1 superior derecho. Vrba (1981).
- Individuo 10. Adulto. KB 5163. C inferior derecho. de Ruiter (2004).
- Individuo 11. Juvenil. KB 5222. M3 superior izquierdo. Vrba (1981).
- Individuo 12. Juvenil. KB 5223. dc, dm1-2 inferiores izquierdos; dm2 inferior derecho; incisivos permanentes inferiores; M1 inferior derecho; M1 inferior izquierdo. Vrba (1981), Grine (1982), Braga y Thackeray (2003).
- Individuo 13. Adulto. KB 5226. M3 inferior izquierdo. Vrba (1981).
- Individuo 14. Adulto. KB 5383. M1 superior derecho. Vrba (1981).
- Individuo 15. Adulto. KB 5389. I1 superior izquierdo. de Ruiter (2004).
- Individuo 16. Juvenil. KB 5503. dm2 inferior derecho. Thackeray et al (2001).
- Posiblemente Individuo 1. KB 5522. Fragmento del eje del húmero izquierdo. Thackeray et al (2005).
- Posiblemente Individuo 1. KB 5524. Cara lingual de una corona molar desgastada, posiblemente M1 o M2. Braga et al (2013).
- Individuo 17. Juvenil. KB 6067 es una porción petrosa del hueso temporal de un individuo juvenil. Su morfología lo relaciona con P. robustus, pero también con StW 53 (Homo gautengensis). Sin embargo, el tamaño de la ventana oval es más parecido al de la de algunos especímenes de Sterkfontein Member 4 (Australpithecus africanus). Si KB 6067 se incluye dentro de P. robustus, esto puede representar una condición evolutivamente menos derivada que la deducida a partir de TM 1517 y otros ejemplares conespecíficos. Braga et al (2013).
Kromdraai E (KE)
Excavaciones a partir de 2014 a cargo del Kromdraai Research Project. Hallazgos pendientes de publicación.
Sterkfontein Miembro 5 (2,18 ma)
Algunos dientes (Granger et al, 2015).
Drimolen (2-1,5 ma)
El análisis de macromamíferos asociados fue realizado por Adams et al (2016).
-
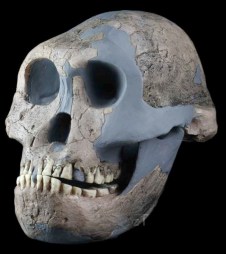
DNH 7 – Paranthropus robustus
DNH 7, Eurydice. 1994. 1,5-2 ma. André Keyser. Cráneo casi completo y mandíbula inferior de una hembra.
- DNH 8 Orfeo. Mandíbula inferior de un macho.
- DNH 44. Mandíbula. Edad al morir 1,70 años (Smith et al, 2015).
- DNH 47. Maxilar y dientes asociados. Edad al morir 0.67-0,77 años (Smith et al, 2015).
- DNH 60. Dientes juveniles asociados con un cráneo.
- DNH 84. Maxilar y dientes asociados. Edad al morir 2,24 años (Smith et al, 2015).
- DNH 101. Molar juvenil aislado.
- DNH 107. Dientes asociados. Edad al morir 4,82 años (Smith et al, 2015).
- DNH 108. Dientes asociados. Edad al morir 5,35-5,53 años (Smith et al, 2015).
Swartkrans, Miembro 1 (2,25-1,7 ma), Miembro 2 (1,7 ma) y Miembro 3 (1 ma)
Es el yacimiento con la muestra más amplia de Paranthropus robustus. Los dientes de Swartkrans son mucho más grandes y por ello se ha sugerido que representan una especie diferenciada: Paranthropus crassidens (Broom 1949, Broom 1950, Howell 1978, Grine 1982, Grine 1985 y Grine 1988). Sin embargo, los restos dentales de Drimolen, de tamaño intermedio, apoyan la hipótesis de una única especie (Keyser et al, 2000; Moggi-Cecchi et al, 2010).
-

KK 48 – Paranthropus robustus
SK 12. Cráneo masculino.
- SK 46. Cráneo masculino.
- SK 48. 1950. Cráneo de un macho (Lockwood et al, 2007). Es el cráneo más completo de P. robustus.
- SK 61. Mandíbula.
- SK 62. Mandíbula. Edad al morir 3,12 años (Smith et al, 2015).
- SK 83 Cráneo masculino.
- SKX 10924. Lague, 2015. Pequeño fragmento distal de húmero. Asignado por Susman, de Ruiter y Brain (2001) a H. erectus.
- Varios dientes descritos por Pickering et al (2016).
- Broom R, Robinson JT (1950) Man contemporaneous with the Sawartkrans ape‐man. Am J Phys Anthropol 8(2), 151-156.
- Robinson JT (1955) Further remarks on the relationship between “Meganthropus” and australopithecines. Am J Phys Anthropol 13(3), 429-445.
- Sillen A (1992) Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated fauna from Swartkrans. J Hum Evol 23(6), 495-516.
- Lee-Thorp JA, van der Merwe NJ, Brain CK (1994) Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis. J Hum Evol 27(4), 361-372.
- Keyser AW (2000) The Drimolen skull: the most complete australopithecine cranium and mandible to date. South African J of Sci 96(4), 189-192.
- Blackwell L, d’Errico F (2008) Early hominid bone tools from Drimolen, South Africa. J Archaeol Sci 35(11), 2880-2894.
- Moggi-Cecchi J, et al (2010) Early hominin dental remains from the Plio-Pleistocene site of Drimolen, South Africa. J Hum Evol 58(5), 374-405.
