a. El pensamiento evolucionista
Las poblaciones humanas están sujetas a las mismas leyes evolutivas que el resto de especies biológicas. El concepto evolucionista es muy antiguo, al menos como idea de cambio que afecta a la Naturaleza. Actualmente somos el resultado de cambios graduales acumulados a lo largo del tiempo y las diferencias inter-individuales y las inter-poblacionales de la especie humana son la base del proceso evolutivo. Esta premisa evolucionista definida por Charles Darwin (1809-1882) se basa en la existencia de variabilidad y en la lucha por la existencia. Pero la idea de cambio no ha sido siempre aceptada. El concepto del Gran Diseño de Aristóteles (384-322 a.C.) supone un único evento de Creación Divina basado en el principio de progresión creativa que explicaría la creciente complejidad de los seres vivos. Nicolaus Copernicus (1473-1543) cuestionó, el Sistema Planetario de Claudius Ptolomeo, (90/100-170 a.C.), sugiriendo que la Tierra gira alrededor del Sol.

Ptolomeus (90/100-170)
En el siglo XVI, James Ussher (1581-1650), arzobispo inglés, encarnó el máximo exponente del fijismo (Las especies no se han modificado desde su creación) al establecer a partir de las Sagradas Escrituras que la Creación Divina tuvo lugar el 20 de Octubre del año 4004 a.C. a las 9:00 horas.
El siglo XVII supone el despertar de la Ciencia. Por primera vez se contrapone el espiritualismo secular con el materialismo que requiere explicación de todos los fenómenos. En esta época René Descartes (1596-1650), pensador y filósofo, establece los principios del método científico en su obra ‘El discurso del método’ (1637). Descartes define cuatro principios básicos para la contrastación de hipótesis que, en esencia, aún están vigentes en nuestra época. Descartes opina que los principios de la Ciencia han de ser tomados de la Filosofía y supone que ‘las cosas que percibimos muy clara y muy distintamente son todas verdaderas’, pero carece del concepto del empirismo que caracteriza a la Ciencia en la actualidad.
Principios del método de Descartes
- No aceptar nunca como verdadera ninguna cosa que no conociese con evidencia que lo era. No comprender nada más que aquello que se presentase tan clara y definitivamente al espíritu que no tuviese ocasión alguna de ponerlo en duda.
- Dividir cada una de las dificultades que se examina en tantas partes como fuera posible y como requiriese su mejor solución.
- Conducir por orden los pensamientos comenzando por los objetos más sencillos y más fáciles de conocer para ascender hasta el conocimiento de los más compuestos.
- Hacer en todo enumeraciones tan completas y revisiones tan generales que se adquiriese la seguridad de no omitir nada.
El siglo XVIII se caracteriza por un enfoque antropocentrista de la realidad. Así, Alexander Pope (1730) considera qie ‘el hombre es el centro’ de todo, pero esta visión llega al máximo con Carolus Linnaeus (1707-1778) y su ‘Systema Naturae’ publicado en 1735. Linneo era fijista en el más amplio sentido, de forma que situó al ser humano en el vértice de la pirámide de la creación.

Linnaeus (1707-1778)
Su gran contribución a la Ciencia fue el sistema dicotómico de clasificación. Linneo consideró que las poblaciones humanas actuales forman parte de dos especies: Homo sylvestris y Homo sapiens, en la que distinguió seis subespecies (H. s. ferus, H. s. monstruosus, H. s. afer, H. s. europeus, H. s. asiaticus, H. s. americanus). A pesar del sesgo creacionista, la taxonomía dicotómica de Linneo estableció un sistema claro de clasificación de los seres vivos que aún perdura en nuestros días.
Buffon (1707-1788), naturalista francés, puede ser considerado el fundador de la Antropología moderna. En su obra ‘Histoire Naturel des animaux’ introduce el concepto de raza y lo relaciona con factores ambientales. Se muestra como un claro evolucionista y establece la antigüedad de la Tierra en 76.000 años. Como naturalista, Buffon es contrario a la sistemática rígida de Linneo. Plantea las relaciones hombre-animal y la variabilidad de las especies. Es el primero en estudiar el grupo humano como especie biológica. Erasmus Darwin (1731-1802), médico rural, poeta y científico, abuelo de Charles Darwin, plantea claramente, junto con Buffon, el concepto de mutabilidad de las especies y cambio orgánico. Aunque no puede explicar dicho proceso, acepta la heredabilidad de los caracteres adquiridos y establece el concepto de selección como motor del cambio.

Lamark (1744-1829)
El primero en proponer una explicación del concepto de evolución orgánica es Jean Baptiste Pierre Antoine de Lamark (1744-1829). La idea de transmutación no era nueva pero sí el mecanismo de acción. En su obra ‘Zoological Philosophy’ (1809) propone un mecanismos de aparición de nuevas especies basado en el uso de órganos y en la herencia de los caracteres adquiridos. También plantea la generación espontánea y la tendencia a la complejidad como mecanismos evolutivos. Lamarck considera que la evolución consiste en el desarrollo orgánico de las especies basándose en tres principios: 1) la generación espontánea de las formas de vida más simples; 2) la evolución de las formas simples hacia las complejas; y 3) la influencia ambiental que tiende a enmascarar esa tendencia a la complejidad. Así, Lamarck definió dos leyes que explicarían el proceso de especiación:
- El uso frecuente y continuo de un órgano lo desarrolla de forma gradual.
- La adquisición o pérdida de un órgano se preserva gradualmente a través de la reproducción.
Pero los principios evolucionistas definidos por Bufon, Darwin y Lamark en el siglo XVII no fueron aceptados por el gran catastrofista George Cuvier en el XVIII. Cuvier no acepta el evolucionismo y mantiene que la existencia de fósiles de especies que no se conocen en la actualidad se explica por la existencia de extinciones masivas en el registro geológico. Las causas de estas extinciones serían diversas: regresión del nivel del mar, enfriamiento global, procesos intensos de predación o competición interespecífica, además de procesos fortuitos como la colisión de asteroides con la Tierra. En esta época preevolutiva de Cuvier cada periodo geológico se consideraba el origen de un nuevo proceso creacionista aislado y habrían existido unos treinta eventos de este tipo.

Tabla 1. Eras, periodos y épocas geológicas y su cronología aproximada.
Desde el punto de vista del catastrofismo de Cuvier, la historia evolutiva de los organismos multicelulares, del llamado periodo Phanerozoico, se divide en tres grandes eras geológicas: Paleozoico (vida ancestral), Mesozoico (vida media) y Cenozoico (vida moderna). Cada una de estas eras se subdividiría en periodos: Cámbrico, Ordovico, Silúrico, Devónico, Carbonífero y Pérmico en el Paleozoico; Triásico, Jurásico y Cretáceo en el Mesozoico; y Terciario y Cuaternario en el Cenozoico. Los periodos más recientes se subdividieron en epocas: Paleoceno, Eoceno, Oligoceno, Mioceno y Plioceno para el terciario; y Pleistoceno y Holoceno para el Cuaternario (Tabla 1).
Esta visión empieza a desaparecer a principios del siglo XIX con el auge del llamado uniformitarismo de Charles Lyel (1797-1875), hijo de un terrateniente inglés. En su obra ‘Principles of Geology’, publicado en 1830, Lyel considera que las fuerzas y procesos geológicos que actúan ahora son responsables de todos los cambios geológicos y se han producido en todas las eras. Estos cambios son en su mayoría graduales (erosión, transporte, sedimentación, etc.) pero ocasionalmente tambiénpueden ser drásticos (meteoritos), pudiendo ser su efecto moderado o catastrófico.
Lo importante es que la respuesta de los ecosistemas en caso de extinción masiva es un rápido periodo de especiación evolutiva que produce la recuperación de la diversidad biológica. El paradigma evolutivo actual se basa en la evolución gradual de las especies pero acepta determinados procesos de extinción rápida que han sido la base de la división geológica del tiempo. Al mismo tiempo, Thomas Robert Malthus (1766-1834), en su obra ‘An essay on the principle of population’ pone de manifiesto la lucha por la existencia como factor limitante del crecimiento de las poblaciones, basándose en dos evidencias: la fertilidad ilimitada de los organismos y la existencia de recursos limitados. Por ello, cada ecosistema tiene una capacidad sustentadora máxima y los organismos deben competir por la supervivencia.
Entrado el siglo XIX, Alfred Russel Wallace (1823-1913) define por primera vez el concepto de selección natural. Wallace viajó al archipiélago Malayo en 1854 para estudiar las aves y los insectos. Basándose en la teoría de Malthus escribió en 1855 un artículo sobre la mutabilidad de las especies. En 1858 publica su obra ‘On the tendency of varieties to depart indefinitely from the original type’ donde plantea su teoría sobre la selección natural. Wallace dedujo la teoría de la supervivencia del más apto a partir de la frase ‘the positive checks to increase’ de la obra de Malthus ‘An eassay on the principle of population’ de 1798.

Darwin (1809-1882)
Casi simultáneamente, Charles Darwin (1809-1882), nieto de Erasmus Darwin, publica su obra ‘On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life’ (1859), donde también presenta su hipótesis sobre la selección natural. Charles Darwin, de familia inglesa acomodada, comienza estudios de Medicina que deja por la carrera de Teología a los 19 años (1828), interesándose al mismo tiempo por las Ciencias Naturales. Se gradúa en 1831, a los 22 años, y se embarca como naturalista en el HMS Beagle, capitaneado por Robert Fitz-roy, el 27 de diciembre de 1831. El viaje dura cinco años y regresa a Inglaterra el 2 de octubre de 1836. Sin embargo, no es hasta 1842 que escribe un borrador de su teoría (12 años antes que Wallace), que corrige en 1844. El 1 de julio de 1858 la Linnean Society de Londres lee en sesión pública las teorías de Wallace y Darwin, pero Darwin no publica hasta 1859, presionado por la obra de Wallace de 1858.
La teoría darwinista de la selección natural se basa en cinco evidencias principales:
- La capacidad reproductora de las especies supera la disponibilidad de recursos.
- Las especies presentan una gran variabilidad de formas.
- Dado que hay más individuos que los que pueden sobrevivir, existe una lucha por la supervivencia.
- Las variaciones favorables sobreviven y pasan a las siguientes generaciones.
- Con el tiempo las variaciones favorables se acumulan y dan lugar a procesos de especiación.
Esta visión darwinista se apoya en diversas evidencias. Por un lado, la selección artificial de animales y plantas genera nuevas variaciones morfológicas y en las poblaciones naturales es posible distinguir variaciones geográficas debidas a aislamiento. Además, el registro paleontológico también muestra un proceso de diversificación y evolución gradual de las especies. Por otro lado, la anatomía comparada reconoce la existencia de caracteres homólogos que teniendo un mismo origen embriológico pueden presentar formas y funciones distintas. En el ámbito embriológico, el desarrollo orgánico muestra etapas que recuerdan estadios evolutivos distintos (la ontogenia recapitula la filogenia), y los órganos vestigiales indican cambios funcionales de dichos órganos.

Charles Darwin, The descent of man – 1871.
En 1871 Charles Darwin publica su obra ‘The descent of man and selection in relation to sex’ donde aborda específicamente la evolución de la especie humana y los factores que la determinan, resaltando la importancia de la adquisición temprana del bipedismo.A pesar de las evidencias actuales de darwinismo, Charles Darwin no consigue explicar el mecanismo de la herencia de los caracteres, sino que adopta la herencia de los caracteres adquiridos para su teoría de la selección natural. De hecho, la teoría mendeliana de la herencia no se publica en Brno por Gregor Mendel (1822-1884) hasta 1865, aunque quedará en el olvido hasta que es redescubierta por Vries, Correns y Tschermak en el año 1900. En 1843 Mendel, que es contemporáneo de Darwin, entra como novicio en el monasterio hasta que obtiene el título de profesor en Viena. En 1856 Mendel inicia experimentos de cruzamientos y desarrollo de híbridos en el guisante (Pisum sativum), hasta que define las leyes de la herencia que llevan su nombre. Mendel se basa en el análisis del número de híbridos producto de cruzamientos entre variedades distintas (color y rugosidad) y en la transmisión generacional de dichos caracteres.
La 1ª Ley de Mendel indica que las variedades genéticas de un carácter no sólo se segregan independientemente sino que se heredan en proporciones constantes de generación en generación.
La 2ª Ley de Mendel pone de manifiesto que caracteres genéticos distintos también se segregan independientemente en el cruzamiento de variedades distintas.

2ª Ley de Mendel – segregación independiente de los caracteres.
Mendel no conocía cuáles eran los mecanismos genéticos generadores de variación (mutación, migración, selección o deriva), pero sus leyes modificaron las ideas preconcebidas sobre la herencia. La teoría del homúnculo, que se remonta a Aristóteles, suponía la existencia de organismos preformados uno dentro de otro y que mediante la reproducción se traspasaban a las siguientes generaciones. Linneo sugirió la teoría de las dos capas, en la que la capa externa de todo organismo procede del padre y da lugar a todo el sistema vascular, mientras que la capa interna derivaría de la madre y generaría el sistema nervioso. La gran aportación de Mendel consistió en determinar que los caracteres se comportan como unidades discretas que se transmites de generación en generación siguiendo modelos matemáticos simples. Actualmente a este tipo de caracteres los denominamos mendelianos y sabemos que existe otros más complejos que no siguen el modelo de herencia sencilla establecido por Mendel.
La teoría de la evolución darwinista tal como la conocemos hoy fue presentada en 1930 por Fisher, Haldane, Wright y Chetverikov. Es lo que se conoce como Neodarwinismo. Sin embargo son Dobzhanski y Mayr en 1937 quienes divulgan realmente el concepto genético del origen de las especies, originándose la Genética de Poblaciones como Ciencia. Desde este punto de vista, la evolución es un proceso que se produce en dos pasos: en primer lugar se origina y redistribuye la variación genética y a continuación la selección natural actúa sobre ella produciendo cambios en las frecuencias alélicas. Las nuevas variantes o alelos se forman por mutación del material genético, constituyendo así nuevas variantes que darán lugar a nuevos fenotipos. Un alelo se localiza físicamente en un gen, que es un fragmento de DNA que codifica una proteína determinada. La selección natural actúa sobre los individuos basándose en los caracteres que presenta y la unidad evolutiva es la población en la que se alteran las frecuencias de los caracteres en fun-ción de su adaptabilidad. El Neodarwinismo de Dobzhanski tiene actualmente diversos problemas, ya que no explica las mutaciones neutras, ni el tempo del proceso evolutivo (microevolución y macroevolución) o las mutaciones sobre genes reguladores. Ya avanzado el siglo XX han aparecido nuevas teorías que completan los mecanismos genéticos y su efecto sobre la evolución de las especies.
b. El marco evolutivo humano
La Biología Humana estudia la evolución espacial y temporal de la especie humana. Originalmente, el término Antropología definió el conjunto de disciplinas que tenían por objeto el estudio del ser humano. Buffon (1707-1778) fue el fundador de la Antropología y estableció tres campos de estudio: 1) análisis morfológico y fisiológico; 2) estudio de las razas humanas; y 3) comparación con otros primates. En 1859 se funda la Societé d’Antropologie de Paris y Brocca (1824-1880) establece los campos de estudio en tres áreas: antropología zoológica (hombre y animales), descriptiva (rasgos humanos) y general (respecto a los demás seres vivos). Valls (1985) distingue entre antropología general (herencia, mutaciones, selección, mezcla, variabilidad, crecimiento, proporciones corporales, diferenciación sexual, etc.), especial o raciología (razas actuales y desapareci-das), genética de poblaciones y métodos antropológicos.
Actualmente distinguimos tres campos de estudio en Biología Humana: 1) Evolución Humana; 2) Genética Humana; y 3) Ecología Humana. Las fuentes de la Biología Humana son diversas. Desde el material esquelético y dentario conservado en los yacimientos paleontológicos o arqueológicos, hasta muestras de células y tejidos, datos médicos, registros personales o demográficos. La obtención de información útil en Biología Humana tiene, sin embargo, limitaciones éticas ya que no es posible estudiar clones o realizar cruzamientos experimentales. Además, la descendencia de la pareja humana es pequeña y el tiempo de gene

Campos de estudio de la Biología Humana
ración es largo, lo que dificulta la obtención de información. Sin embargo, estas deficiencias se ven compensadas por el elevado número de poblaciones y efectivos muestrales naturales que existen, de forma que siempre será posible buscar en las poblaciones naturales muestras que se adapten al estudio que queremos reali-zar. Por ejemplo, podemos estudiar gemelos que nos permiten realizar inferencias sobre la influencia del ambiente sobre determinados caracteres genéticos.
La evolución de las poblaciones humanas (descendencia con modificación) depende de la capacidad reproductora, que generalmente se mide por el tamaño poblacional, y de la supervivencia diferencial de los individuos. Así, el proceso evolutivo de las especies se puede estudiar mediante el análisis tanto de los cambios morfológicos como de las frecuencias alélicas de los caracteres. Además, también es posible analizar la variación del número de taxa (evolución taxonómica). En los últimos seis millones de años de evolución de los homininos, el número de taxa ha variado en función de las condiciones ambientales y los procesos de adaptación ecológica.
Como hemos visto, la división de la escala temporal en eras y periodos se basa en grandes procesos de extinción de especies y desaparición de taxones. Las causas de dichas extinciones masivas son diversas e incluyen la regresión del nivel del mar, el enfriamiento global, la predación de especies o la competición interespecífica. La respuesta biótica a las extinciones es la radiación adaptativa favorecida por la existencia de hábitats nuevos sin competencia. Esto introduce un factor aleatorio de la evolución, incluso oportunista, que puede originar procesos de macroevolución.

Las 5 principales extinciones masivas del registo fosil.
La aparición de una nueva especie se produce por aislamiento reproductivo. Una especie natural es un grupo de organismos potencialmente capaces de reproducirse entre si (criterio biológico), pero no con otras especies. Por tanto, se trata de un sistema genéticamente cerrado, a diferencia de las subespecies o razas que son sistemas abiertos. La especiación transforma un grupo poblacional abierto en cerrado por aislamiento genético, originando especies simpátridas si sus rangos de dispersión geográfica se superponen, alopátridas si no se superponen sus áreas de distribución y alocrónicas si lo que no se superpone son sus rangos de distribución temporal, por lo que se dice que son cronoespecies. El éxito evolutivo de una nueva especie se debe a su adaptabilidad a nuevas condiciones ambientales. Si la nueva especie ocupa un hábitat nuevo, sin competencia, se favorece su adaptación o incluso su diversificación, pudiéndose originar una radiación adaptativa, como es el caso de la radiación de los Primates tras la extinción de los Dinosaurios hace unos 65 millones de años (ma). En cambio, si la nueva especie se adapta a un hábitat ya ocupado, se produce una exclusión competitiva que culmina con la eliminación de uno de los grupos, ya sea porque evita la competencia, abandona el hábitat, se readapta o se extingue al no poder competir.
El proceso de especiación se pone de manifiesto por la evolución de caracteres adaptativos nuevos, que se originen por la especialización a nuevas condiciones de vida. Estos caracteres se denominan especializados o derivados y se diferencian de los caracteres primitivos que suelen ser no especializados o generalizados. Se denomina polaridad de un caracter a su clasificación respecto a si es primitivo o derivado. Los conceptos de primitivo y generalizado o derivado y especializado no tienen por qué coincidir. Por ejemplo, la mano humana con cinco dedos es un carácter primitivo ya que está presente en los primeros primates hace 65 ma, pero aunque se trata de una carácter generalizado (no especializado), determinados aspectos de la mano humana (como la precisión del movimiento del pulgar) constituyen verdaderas especializaciones evolutivas. Atendiendo a su origen, los caracteres evolutivos pueden clasificarse también en homólogos, aquellos que derivan de un antecesor por descendencia común, o análogos, aquellos que son únicos en las formas descendientes y no están presentes en el ancestro. El análisis del origen evolutivo de los caracteres adaptativos es lo que nos permite establecer filogenias o relaciones de antecesor-descendiente entre especies.
c. Principios de clasificación filogenética
La filogenia de un grupo de especies se basa en la comparación de caracteres evolutivos entre ellas. Existen dos escuelas principales de clasificación. La Fenética o taxonomía numérica, establece las relaciones entre especies mediante las similitudes anatómicas generales entre ellas, enfatizando la adaptación, que es una diferencia de grado a expensas de la filogenia real del grupo ya que algunas similitudes morfológicas pueden deberse a convergencia evolutiva o a evolución paralela sin que los taxones tengan un origen filogenético próximo. La escuela del Cladismo o sistemática filogenética, en cambio, prioriza la filogenia estableciendo diferencias de clado sin considerar las analogías ya que son caracteres que no derivan de un ancestro común.

Clasificación de los caracteres anatómicos según las teorías gradística (primitivo, derivado) y cladística (plesiomorfía, apomorfía).
En un término medio entre fenética y cladismo tendríamos la denominada escuela evolutiva. Si la evolución progresa de forma regular, fenética y cladismo coinciden. Los paleontólogos y morfólogos suelen utilizar aproximaciones fenéticas basadas en caracteres morfológicos, mientras que la genética y la biología molecular aplican más los conceptos cladistas. El cladismo considera que sólo los caracteres homólogos y derivados pueden definir filogenias, especialmente aquellos que son compartidos entre dos o más taxones. El problema está en definir cuáles son estos caracteres. En el ámbito terminológico, podemos distinguir tres tipos de caracteres evolutivos: apomorfías, plesiomorfías y homoplasias. Las apomorfías son caracteres derivados; pueden ser autapomorfías si son exclusivos de un taxón o sinapomorfías si son compartidos con otros taxones. Las plesiomorfías son caracteres primitivos que derivan por descendencia de un taxón ancestral, y se denominan simplesiomorfías si son caracteres primitivos compartidos con otros taxones. Las homoplasias son caracteres análogos debidos a convergencia adaptativa o evolución paralela que no permiten establecer relaciones filogenéticas.
Los caracteres evolutivos y su naturaleza determinan el tipo de grupo evolutivo que forman los taxones que analizamos. Así, los grupos pueden ser monofiléticos si comparten un carácter homólogo derivado ancestral al clado (simplesiomorfía), polifiléticos si miembros de diferentes linajes convergen en una adaptación similar (homoplasias) y parafiléticos si contienen subgrupos de descendientes de un ancestro único.

Ejemplos de grupos taxonómicos monofilético (reptiles y aves), parafilético (reptiles) y polifilético (aves y mamíferos).
El establecimiento de filogenias para caracteres morfológicos requiere separar las homologías de las homoplasias para establecer grupos monofiléticos respecto a taxones externos de comparación (outgroups) con los que no comparten homologías. En cambio el establecimiento de filogenias a partir de caracteres moleculares (sistemática molecular) se basa en la obtención de datos moleculares (secuencias de DNA, hibridación, electroforesis, reacciones inmunológicas, etc.) y su análisis específico mediante técnicas basadas generalmente en el cálculo de distancias. La distancia genética es una medida fenética de diferencia o similitud general entre taxones. Por ello, los cladistas rechazan estos métodos y sólo aceptan los que se basan en el análisis de secuencias de DNA por ser rasgos en los que la determinación de su estado se realiza de forma directa (presencia o ausencia de un determinado nucleótido).
La sistemática molecular se basa en el hecho de que cuando dos especies divergen las mutaciones se acumulan independientemente en el DNA de los linajes descendientes y su análisispermite establecer relaciones de afinidad entre especies. Inicialmente los biólogos moleculares pensaron que se trataba de un método superior a los tradicionales. Primero porque la información deriva de los propios genes de las especies (portadores del registro fundamental del cambio evolutivo). Segundo, porque se suponía que la información molecular no estaba afectada por la convergencia evolutiva (homoplasia), ya que esta se produce por adaptación ambiental y la mayoría de mutaciones son neutras y, por tanto, invisibles a la selección. Y tercero, porque la evolución molecular se supuso constante y regular en el tiempo, a diferencia de la morfológica que es siempre errática. Todo ello llevó a definir el concepto de reloj evolutivo molecular que permitiría, no sólo definir el orden de las ramificaciones de especies relacionadas, sino también el momento de la divergencia de cada linaje y la longitud de las ramas.
La sistemática molecular y la morfolólogica a menudo proporcionan resultados contradictorios y sólo si se comparan con filogenias conocidas se podrá comprobar su validez. Existen diversas limitaciones de la sistemática molecular. Por un lado, la dinámica de las mutaciones es compleja y no todas las regiones moleculares son igualmente susceptibles al cambio. Además, la convergencia es posible ya que hay regiones que son susceptibles a un determinado tipo de cambio y existen mutaciones más probables que otras (mutaciones recurrentes). Distintos genes pueden mutar con tasas diferentes haciendo que el reloj molecular no sea constante, o incluso un mismo gen puede presentar variaciones en la tasa de mutación en dos linajes distintos o en dos momentos distintos, por lo que la noción de un reloj global desaparece, aunque pueden existir relojes locales que sí son útiles. Además, algunas mutaciones pueden desaparecer por retromutación y la tasa de divergencia genética puede no reflejar la tasa de divergencia de las especies (árbol de genes vesus árbol de especies). La filogenia verdadera es la de las especies, no la de los genes, ya que los genes pueden divergir antes de la especiación.

Separación de los procesos de divergencia de especies y de los genes.
El concepto de homología para las secuencias genéticas es complejo y requiere algunas consideraciones. El término homología se refiere al ancestro común mientras la similitud se refiere a la apariencia de las secuencias derivada, por lo que no hay que hablar de homología cuando nos referimos a similitud entre secuencias. Genes que tengan poca similitud en su secuencia pueden ser homólogos si mantienen su estructura tridimensional que le confiere una funcionalidad específica. La duplicación de genes (frecuente en muchos organismos) complica la cuestión ya que cada copia puede acumular variantes de forma independiente. En genes no duplicados (de copia única) se aplica el término ortología, que es comparable a la homología, y en genes duplicados se habla de paralogía en una familia de genes. Una tercera forma de homología es la xenología, que ocurre cuando se producen saltos de las barreras entre especies (transmisión horizontal de genes), en general por acción de virus que transfieren un gen de otra especie a un huesped. La xenología se detecta cuando en dos especies no emparentadas aparecen secuencias que son similares. Este tipos de homología parcial no ocurre en los caracteres morfológicos.
Paradójicamente una de las ventajas de la sistemática morfológica es el carácter errático de su tempo evolutivo. Un claro ejemplo es la radiación evolutiva, que genera muchos cambios morfológicos en un corto periodo de tiempo. Un reloj mutacional constante no sería capaz de registrar todos los cambios si la radiación se produjo muy atrás en el tiempo. Un reloj rápido sí recogería el evento pero sería borrado posteriormente por el alud mutacional. En cambio, los cambios morfológicos persisten en los linajes posteriores (por ejemplo, la radiación de los mamíferos placentarios hace 100 ma). Por contra, una gran ventaja de la sistemática molecular es el gran volumen de información que puede procesar, pudiendo mezclar en un mismo análisis posiciones con tasas de mutación bajas (DNA ribosómico) y altas (DNA mitocondrial).
d. Técnicas de datación
La visión evolutiva introducida por Darwin en The Origin of Species considera que las especies extinguidas representan formas adaptativamente inferiores, menos adaptadas que sus sucesores. El hecho de la extinción de especies, incluso masivamente, ya había sido demostrado antes que Darwin por el anatomista francés Georges Cuvier, a finales del siglo XVIII. Cuvier demostró que los huesos de mamut descubiertos en la depresión geológica de París eran distintos a los del elefante actual y que eran, por tanto, representantes de una especie distinta que se extinguió hace mucho tiempo. Cuvier identificó lo que creyó que eran periodos de extinción masiva de numerosas especies, o catástrofes, acaecidas en cortos periodos de tiempo. Estas observaciones inspiraron múltiples trabajos geológicos durante principios del siglo XIX, en los que se identificaron intervalos de aparente cambio en la historia evolutiva de las especies y se definieron límites entre lo que se llamó periodos geológicos.
Desde el punto de vista del Catastrofismo de Cuvier, la historia evolutiva de los organismos multicelulares, del llamado periodo Phanerozoico, se divide en tres grandes eras geológicas: Paleozoico (vida ancestral), Mesozoico (vida media) y Cenozoico (vida moderna) (Lewin 1998, p. 69) y cada una de ellas se subdividiría en periodos geológicos que se consideraba el origen de un nuevo proceso creacionista aislado (habrían existido unos 30 eventos de este tipo según la teoría catastrofista). Esta visión cambia radicalmente con el auge del llamado Uniformitarismo de Charles Lyel (Principles of Geology, 1830) que considera que las fuerzas y procesos geológicos que actúan ahora son responsables de todos los cambios geológicos y se han producido en todas las eras geológicas. El paradigma evolutivo actual se basa pues en la evolución gradual de las especies pero acepta determinados procesos de extinción rápida (catastrófica) que han sido la base de la división geológica del tiempo. La reconstrucción de los patrones evolutivos de caracteres anatómicos o del comportamiento requieren un marco temporal preciso ya que de otra forma no sería posible reconstruir el tempo, la sucesión, en que se produjeron los cambios evolutivos. Actualmente existen distintos métodos de datación que cubren, con algunos huecos, desde hace unos 1.000 años hasta varios billones. La Paleoantropología se centra, sin embargo, en los últimos 10 millones de años.
Los métodos de datación pueden ser directos o indirectos. Los métodos directos datan el propio fósil del que queremos saber la antigüedad, mientras que los indirectos se aplican sobre materiales con una antigüdad supuestamente igual a la del material que queremos datar. No todos los materiales son susceptibles de dataciones directas ya que las técnicas que se utilizan sólo son aplicables a determinados materiales. Algunos métodos, como la datación por carbono-14 (C14) y la resonancia electrónica de spin (ESR) pueden ser aplicados directamente sobre dientes y fósiles recientes y también sobre pigmentos y pinturas en cuevas. La datación por termoluminiscencia se puede aplicar a vasijas antiguas, lascas y granos de tierra. Por lo general, las técnicas directas son destructivas y por tanto no son apropiadas para materiales de gran valor. En la práctica, por tanto, los métodos indirectos son los más utilizados. En este caso, la datación se aplica a un fósil o artefacto que está en asociación con el que queremos datar.
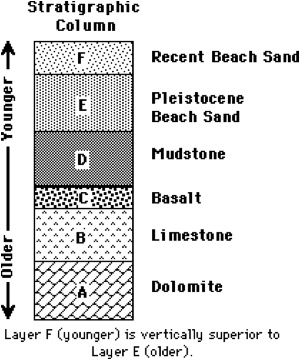
Principio de Superposición Estratigráfica: los estratos más antiguos están debajo de los más recientes.
La datación indirecta se basa en el principio arqueológico de superposición estratigráfica que supone, salvo excepciones, que los niveles estratigráficos más antiguos se encuentran debajo de los más recientes y, por extensión, dos artefactos situados en el mismo nivel estratigráfico son contemporáneos. Así, podemos determinar la edad aproximada de un fósil datando los niveles inmediatamente inferior y superior al que se encuentra dicho fósil. De esta forma, fijamos un rango de antigüedad en el que se encuentra el material que queremos datar. Además de directos o indirectos, los métodos de datación pueden ser relativos, si establecen la antigüedad de un material por comparación con otro de otra localidad o yacimiento, o absolutos, si proporcionan una medida física de la edad del material.
Las técnica de datación relativa más utilizadas son el paleomagnetismo, la dendrocronología, la seriación estratigráfica y la correlación faunística. Dado que la escala geológica del tiempo se basa en la historia de la vida sobre la Tierra y en los principales cambios de las poblaciones fósiles, como la aparición o desaparición de grupos, podemos aplicar secuencias estratigráficas conocidas (como la historia evolutiva de los cerdos, elefantes o caballos) para establecer dataciones de fósiles asociados a un determinado hallazgo faunístico. Estos grupos de referencia, actualmente ampliamente conocidos, muestran cambios evolutivos y tendencias específicas, por lo que un correcto análisis estratigráfico y faunístico de un yacimiento permite realizar dataciones bastante precisas.
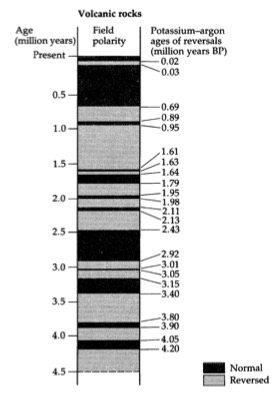
Fluctuaciones de las secuencias paleomagnéticas normal e inversa de los últimos 4,5 ma.
El paleomagnetismo se basa en el hecho de que el eje magnético de la Tierra no está fijo sino que se invierte periódicamente. Actualmente nos encontramos en un periodo de polaridad normal ya que el Norte magnético coincide aproximadamente con el Norte geográfico o verdadero ( la declinación magnética respecto al Norte verdadero es de unos 4º hacia el Oeste). Durante las inversiones del campo magnético, el Norte magnético se sitúa al Sur geográfico. En el momento de la formación de las rocas, especialmente las volcánicas mediante erupciones, la orientación del campo magnético queda registrada por las partículas que contienen hierro, ya que las órbitas de giro de los electrones se ven afectadas por el campo magnético imperante en cada momento. Esto permite establecer secuencias periódicas de normalidad o inversión que ayudan a fijar la edad de un fósil en un intervalo específico. Sin embargo, es un método poco preciso, que se suele utilizar en combinación con otras técnicas, en particular las técnicas de datación absoluta (Lewin 1998, p. 86).

Principio de seriación estratigráfica entre zonas geográficas distintas.
La dendrocronología se basa en la superposición de los anillos de crecimiento de los áboles, que sigue un patrón que depende de las fluctuaciones estacionales de las condiciones de temperatura y lluvia que determinan un patrón de bandas de crecimiento de los anillos de los árboles que es específico de cada zona. La comparación de los anillos permite determinar si una secuencia es anterior o posterior a otra, pero no proporciona una antigüedad específica.
La seriación estratigráfica permite establecer patrones de sucesión de estratos entre regiones distintas, por lo que se puede determinar si su antigüedad es mayor o menor que la serie descrita en otra zona. Las relaciones entre estratos se establece a partir de sus características geológicas y por correlación faunñistica. Los noveles que tienen la misma composición y contienen materiales fósiles similares se asocian para reconstruir la historia sedimentaria de una región.
La mayoría de los métodos directos son radiométricos y se basan en dos principios generales: 1) requieren una puesta en marcha (el reloj se pone a cero) y 2) registran el tiempo mediante el decaimiento constante de un compuesto radioactivo.
Datación por Potasio/Argón (40K/40Ar) y Argón/Argón (39Ar/40Ar)

Técnica de datación absoluta Potasio/Argón.
El 40K (Potasio-40) es un isótopo radioacti-vo (respecto al isótopo estable 39K) que constituye el 0.01% de todo el K disponible y que se descompone progresivamente en 40Ar (Argón-40), que es un gas inerte. Las rocas que contienen K (como las rocas volcánicas) acumulan 40Ar en su estructura cristalina de forma progresiva. La elevada temperatura durante la formación de la roca elimina el Ar y otros gases por lo que la concentración inicial de Ar es cero (el reloj se pone en marcha). La concentración de Ar de una roca volcánica depende del tiempo transcurrido y de la concentración inicial de K. El cálculo de la edad a partir de la concentración de 40Ar se hace mediante comparación con mediciones sobre rocas ricas en potasio como los feldespatos. Un problema importante a tener en cuenta en la datación 40K/40Ar es la contaminación de las muestras con otras de periodos distintos, lo que es frecuente en las erupciones volcánicas. Esta técnica se utilizó en 1960 en la datación del fósil de Zinjanthropus encontrado por Mary Leakey (esposa de Louis Leakey) en 1959 en los niveles de Olduvai Gorge, proporcionando una fecha de 1,75 ma.
Dos avances metodológicos han hecho que la técnica de 40K/40Ar sea de gran fiabilidad. Uno de ellos consiste en que desde los años 60 es posible medir la concentración de ambos isótopos en la misma muestra en lugar de muestras separadas. La técnica consiste en transformar el 39K en 39Ar por radiación de neutrones; después se liberan ambos isótopos de Ar en forma gaseosa por calentamiento con laser y se mide su concentración por cromatografía de gases. La cantidfad de 39Ar es una estima del 40K original de la roca y el 40Ar mide el decaimiento del 40K desde la formación de la roca. Esta técnica se conoce como 39Ar/40Ar. El otro avance, desarrollado en los años 80, es que es posible aplicar esta técnica a cristales aislados de ceniza volcánica. La técnica se conoce como fusión por láser de cristales aislados y permite evitar los problema de la contaminación. El límite de esta técnica es aproximadamente 500.000 años para los materiales más recientes, aunque si se trata de rocas con minerales ricos en K puede llagar hasta 10.000 años. No hay un límite superior efectivo.
Datación por Trazas de Fisión (“Fision Track”)

Trazas de fisión en un cristal de una roca.
Se usa habitualmente junto con el método de Potasio/Argón. El cristal natural contiene el isótopo radioactivo 238U que decae por fisión violenta, lo cual deja una huella o traza sobre el cristal. A mayor tiempo desde la erupción, momento en que se forma el cristal, mayor número de nuevas trazas formadas. El método de preparación de la muestra es lento y el contaje de trazas no es siempre fiable. Pueden datarse mediante esta técnica materiales recientes (algunos miles de años) pero en la práctica la datación es más fiable cuanto mayor la antigüedad del material.
Los dos métodos descritos (Potasio/Argon y Trazas de Fisión) se basan en la existencia de erupciones volcánicas, que fueron frecuentes en África durante la evolución de los hominini, pero no tanto en Eurasia durante los periodos de ocupación humana.
Datación Radiocarbónica (14C)
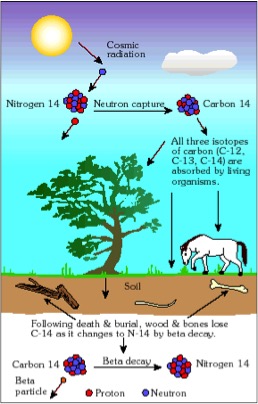
Ciclo del Carbono en los organismos.
Es el método más conocido pero con un rango temporal de aplicación pequeño. La mayoría de Carbono atmosférico del CO2 se encuentra como isótopo 12C estable y una pequeña parte como 14C, un isótopo radioactivo que decae con rapidez. La relación entre ambos isótopos en los vegetales y en los animales refleja la relación atmosférica del momento, ya que el Carbono de los vegetales forma los tejidos de los animales.
Tras la muerte del organismo, el equilibrio entre los isótopos se deshace a medida que el 14C decae. La tasa de transformación de 14C en 12C decrece a medida que la cantidad de 14C disminuye, lo que constituye la base del reloj. En principio cualquier material puede ser analizado mediante esta técnica, pero en la práctica en algunos materiales el 14C decae con mayor rapidez que en otros. El material más apropiado para la datación con esta técnica absoluta es el carbón procedente de cenizas, pero también se han datado radiocarbonicamente pigmentos y huesos. La contaminación de la muestra también es un problema ya que la inclusión de carbones recientes puede variar la datación. La utilización de la espectrometría de masas por aceleración (Accelerator Mass Espectrometry) permite reducir la cantidad de muestra necesaria y amplía el rango de dataciones desde varios cientos hasta unos 100.000 años. El método de Carbono-14 debe ser corregido dado que en la atmósfera la concentración de isótopos del Carbono no ha sido siempre la misma. Las correcciones se hacen utilizando las lecturas radiocarbónicas de los anillos de crecimiento de los árboles, que permiten correlacionar las dataciones no corregidas con la antigüedad real de los anillos.
Datación por series de Uranio/Torio
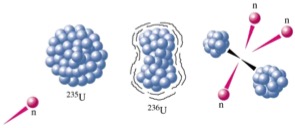
Se basa en que los radioisótopos 238U, 235U y 232To decaen progresivamente hasta formar isótopos estables de Plomo. El método de data-ción por Uranio depende de su solubilidad. A medida que el Uranio decae, sus productos derivados tienden a precipitar de forma que la cantidad de 230To en el sedimento marino de-pende de su antigüedad. No es aplicable a val-vas ya que incorporan Uranio soluble del medio acuático. La serie del Torio sí es aplica-ble a los carbonatos ya que el Torio (Th) y el Protactinio (Pa) son ambos insolubles y el Ura-nio incorporado a las valvas decae formando dichos productos. Las relaciones 230Th/234U (llega hasta 350.000 años) y 232Pa/235U (hasta 150.000) son las más útiles. Se puede aplicar a estalactitas de cuevas y a valvas de yacimientos arqueológicos.
Datación por Racemización de Aminoácidos
Este método se basa en la lenta transformación de la conformación de los aminoácidos (aa), que es levógira en organismos vivos pero que tras su muerte se convierten progresivamente en dextrógira por racemización, formando una mezcla de formas levógiras (L) y dextrógiras (D). La relación entre formas D y formas L depende del tiempo transcurrido. Todos los aminoácidos (excepto la glicina) sufren la racemización, pero el ac. aspártico es particularmente útil. Tiene una vida media de unos 15.000 años.

Configuración de formas L y D de una aa como resultado del proceso de racemización.
La racemización es un proceso que depende de la temperatura (es más rápido a mayor temperatura) por lo que es necesario calibrar la relación D/L y si disponemos de dataciones radiocarbónicas podemos convertir el índice en una datación absoluta. El rango efectivo de la racemización está entre 5.000 y 100.000 años. Esta técnica se ha aplicado a un fragmento de fémur humano encontrado en Abri Pataud, en la Dordogne francesa. Se obtuvo un índice D/L=0.123 y por correlación con dataciones radiocarbónicas se obtuvo una fecha de 16.000 B.P. que estaba de acuerdo con datos arqueológicos y culturales del yacimiento. Sin embargo, recientemente se ha utilizado la relación D/L como un indicador de conservación del DNA y de preservación de la muestra. Esto podría ser un inconveniente ya que no en todos los yacimientos las condiciones de enterramiento son las mismas, pudiendo afectar al proceso de racemización.
Termoluminiscencia y Resonancia Electrónica de Spin

Liberación de energía residual mediante termoluminiscencia o resonancia magnética en materiales cristalinos.
Ambos métodos se basan en el principio de que los electrones de algunos minerales se excitan al ser irradiados por radioisótopos de Uranio, Torio y Potasio (isótopos que están presentes de forma natural en el suelo y en los rayos cósmicos). Los rayos radioactivos chocan con los electrones, cargados negativamente, haciéndolos saltar y dejando una carga positiva en el átomo. Algunos electrones liberados pueden volver a ocupar los “huecos” positivos pero otros son “atrapados” por zonas defectuosas del cristal mineral y se mantienen a un nivel intermedio. La exposición al calor sitúa los electrones atrapados de nuevo en su lugar y se reinicia el proceso. El número de electrones atrapados en niveles energéticos intermedios es proporcional al tiempo transucrrido desde la exposición del material a la acción de materiales radioactivos.
En la técnica de la termoluminiscencia el material que se quiere datar es calentado progresivamente hasta que libera los electrones atrapados, que al volver a su posición original liberan fotones (luz) que pueden ser detectados por instrumentos de precisión. La resonancia electrónica mide directamente los electrones atrapados ya que estos actúan como diminutos imanes que se orientan al ser expuestos a un campo magnético fuerte. La energía de microondas hace cambiar la orientación de los electrones, proporcionando una señal característica cuya intensidad es proporcional al número de electrones atrapados. Ambas técnicas pueden proporcionar dataciones entre varios miles y 1 millón de años, y se sitúan entre los límites del Carbono-14 y el Potasio/Argon. Esta técnica se aplicó a los fósiles de Neandertal y humanos modernos de Próximo Oriente, resultando que los modernos tenían 100.000 años de antigüedad en lugar de los 50.000 que se creía utilizando otras técnicas, mientras que los Neandertales de la misma zona tienen 60.000 años, por lo que no podían ser sus ancestros.
Datación por hidratación de la obsidiana
Es un método simple pero no muy preciso. Se basa en la capacidad de hidratación de la obsidiana. Una vez expuesta una superficie (por ejemplo durante la fabricación de utensi-lios líticos de obsidiana) el proceso de hidrata-ción empieza y la banda de hidratación (observable en un corte en lámina fina) será más gruesa cuanto más tiempo haya transcurrido. La ecuación para determinar la edad en años debe determinarse para cada zona y periodo cronológico t depende del grosor de hidratación en micras. Pero en función de la composición de la obsidiana la tasa de hidratación puede variar hasta tres o más veces su valor.
Secuencias arqueológicas: evolución cultural
La Cultura humana, desde su aparición, también ha sido objeto de datación y se han desarrollado diversas técnicas de estudio. Se ha intentado establecer un paralelismo entre secuencias estratigráficas y secuencias culturales. El investigador francés Gabriel de Mortillet (1821-1898), profesor de la Escuela de Antropología de París en 1978, cuestionó la división de las culturas de la Edad de la Piedra que había realizado Lartet en base a los animales que habían constituido la caza principal de los seres humanos de cada época. Así, Lartet distinguía entre las fases del mamut, el oso cavernario, el caballo salvaje y finalmente el reno. En 1969 Mortillet propuso una nueva división de la Edad de la Piedra en base a los tipos característicos de sus instrumentos líticos, dividiendo las culturas en tres grupos: prechelense (fase anterior al hacha de mano con utensilios rudimentarios), chelense (hacha maciza a modo de martillo de mano con ambas superficies toscamente retocadas) y Achelense (hachas de mano bifaciales con superficies finamente retocadas). Esta clasificación fue revisada por el abate Breuil que unificó el chelense y el achelense en una sola categoría y denominó abbevilliense al prechelense. Otras modificaciones posteriores fueron introducidas hasta llegar a la clasificación más generalizada actualmente. La secuencia cultural básica que puede aceptarse es olduvaiense (modo 1), achelense (modo 2), musteriense (modo 3 y auriñaciense (modo 4), con sus variaciones solutrense, magdaleniense y azilliense asociadas en lineas generales a determinadas formas de la línea evolutiva de los humanos.

Modos tipológicos 1 olduvaiense (arriba izquierda), 2 achelense (arriba), 3 mousteriense (centro) y 4 auriñaciense (abajo).
Estas clasificaciones de la industria lítica se basan en el estudio de sus atributos característicos, ya sean cuantitativos (métricas) o cualitativos, del artefacto considerado. La tipología se encarga del estudio de los atributos de los artefactos líticos con el fin de establecer tipos morfológicos caracterizados por la asociación temporal y/o espacial de determinados atributos. En especial, las series temporales han tenido una especial importancia en las clasificaciones tipológicas y se ha tendido a delimitar rangos evolutivos de la tecnología lítica. Una serie temporal determina el momento en que surge un complejo tipológico, su expansión y su declive hasta que es sustituido por otro.
La seriación de tipos permite establecer cronologías que pueden ser utilizadas para datar determinados yacimientos en función de la industria lítica que contienen. Es una técnica en general poco fiable ya que no están claros los límites culturales de las tipologías, que no dejan de ser subjetivas. Históricamente se asoció cada complejo tipológico con un determinado estadio evolutivo de las poblaciones humanas, pero actualmente el panorama es más complejo y en Arqueología se tiende a considerar el argumento de funcionalidad por encima del de tipología. Es cierto que la tipología también asocia los conceptos de forma y función (rascadores, puntas, cuchillos, buriles, etc.) pero son cada día más los arqueólogos que consideran que la forma es también consecuencia del desarrollo tecnológico y que la función no depende de aspectos tipológicos. Además, la superposición y alternancia entre tipos, especialmente en épocas recientes, es muy grande. Sí es cierto, sin embargo, que determinadas cultural líticas están asociadas preferentemente a grupos humanos concretos. Por ejemplo, la industria musteriense con la técnica Levallois está claramente asociada a los Neandertales del Paleolítico Medio en Europa.
e. Paleoclimatología
En determinados casos es posible situar cronológicamente un fósil si tenemos información sobre aspectos paleoclimáticos del nivel estratigráfico en que se encontró. Dicha información proviene de múltiples fuentes (palinología, análisis faunístico, etc.) pero actualmente se establecen secuencias climáticas en base a la proporción de isótopos del Oxígeno en los caparazones de foraminíferos de niveles estratigráficos marinos profundos o en los glaciares, indicadores de la temperatura preponderante y del volumen total del hielo sobre la tierra.

La proporción entre los isótopos 16O y 18O del Oxígeno del agua permite determinar las fluctuaciones climáticas de épocas pasadas.
La técnica se basa en la cuantificación de la proporción entre el 18O y el 16O, que en los foraminíferos refleja la proporción existente en el medio marino en el que se formaron (a partir del CO2 disponible). El 18O es un isótopo del Oxígeno que, por ser más pesado que el 16O, tiene mayor resistencia a la evaporación y tiende a precipitar con mayor facilidad. Por ello, el agua evaporada contiene más 16O que 18O, que tiende a quedar atrás. En periodos fríos la formación de casquetes glaciares retiene el 16O en los continentes, donde aumenta su concentración relativa. El paso a un periodo cálido devuelve el 16O al mar lo que hace disminuir la concentración relativa de 18O. Por ello, la relación δ18/16O será menor en las masas de hielo en periodos fríos que en periodos cálidos.
Esta y otras técnicas han permitido reconstruir las fluctuaciones climáticas de los últimos 65 millones de años. Podemos dividir el Cenozoico en 4 grandes periodos:
- Entre 65 y 35 millones de años (durante el Paleoceno y el Eoceno) el clima era en general cálido y seco con pocasfluctuaciones, con un pico cálido al principio del Eoceno y un progresivo enfriamiento hacia final del Eoceno.
- Entre 35 y 15 millones de años (Oligoceno y parte del Mioceno) las temperaturas eran superiores a las necesarias para formar casquetes polares antárticos permanentes, con un pico cálido al principio del Mioceno.
- Hace unos 15 millones de años se formó un casquete polar en la Antártida y la temperatura media del mar era de unos 5ºC por encima de la actual. Estas condiciones se mantuvieron hasta hace unos 3 millones de años con oscilaciones de periodos fríos, pero en conjunto se mantuvo una climatología preglacial.
- Hace unos 3,2 ma se iniciaron periodos de unos 100.000 años de amplias fluctuaciones climáticas como respuesta a las glaciaciones del hemisferio Norte. La escala de estas glaciaciones aumentó hace 2,4 ma de nuevo hace 0,8 ma. Los periodos glaciales de los últimos 3,2 millones de años se caracterizan por clima frío, con hielo continental, bajo nivel del mar y gran estacionalidad. Los interglaciales, en cambio, estaban relativamente libres de hielo continental, eran menos estacionales y más cortos. En los últimos 800.000 años las fluctuaciones climáticas se intensificaton como consecuencia de factores astronómicos como la excentricidad de la órbita terrestre, oblicuidad y precesión del eje terrestre.

Fluctuaciones climáticas de los últimos 65 millones de años estimadas a partir de los isótopos estables de Oxígeno.
Las fluctuaciones climáticas están asociadas a fluctuaciones biológicas. Así, existen diferencias entre especies adaptadas al frío y al calor. Algunas especies son tolerantes a los cambios ambientales (especies euribiómicas) mientras que otras son especialistas con poca tolerancia (especies estenobiómicas), lo que las convierte en índices representativos de periodos climáticos concretos. El concepto de fósil índice se utiliza ampliamente en arqueología como indicador de un periodo cronológico determinado. Por ejemplo, la especie Mymomis savinii se ha utilizado como índice en toda Europa para caracterizar niveles anteriores a 500.000 años ya que se trata de un pequeño roedor que se extingue completamente a partir de esta fecha.
