La transición de Australopithecus a Homo es un tema de debate, tanto por la ausencia de fósiles entre 2,5 y 3 ma como por la gran diversidad de los restos disponibles. Es posible que los fósiles más antiguos asignados a Homo (Early Homo) representen diversos linajes. Las evidencias anatómicas de los primeros representantes del género Homo retienen plesiomorfías presentes en Australopithecus, en particular en la dentición y el aparato locomotor, lo que les aproximaría más a los australopitecinos que a Homo sapiens (Wood, 2009). Los fósiles más antiguos atribuibles al género Homo aparecen aproximadamente hace 2,8 ma pero no es fácil definir qué apomorfías a este género. Se suele argumentar que una mayor calidad de la dieta, incluyendo carne, y una mayor flexibilidad de comportamientos en un periodo de cambios climáticos, son los factores principales que explican el origen y la evolución de Homo. Los rasgos que se hacen evidentes a lo largo de la evolución del linaje son 1) el aumento de tamaño del cerebro y del cuerpo en general, 2) la disminución del dimorfismo sexual, 3) un desarrollo lento con alargamiento del periodo infantil, 4) y una creciente cooperación entre los individuos. Pero estas características no aparecen de golpe y los hallazgos fósiles muestran una gran variabilidad de tamaños y formas. Los restos hallados por los Leakey entre 1960-1964 se describieron inicialmente como un australopitecino grácil. Finalmente, Leakey, Tobias y Napier los incluyeron en una nueva especie: Homo habilis. Sin embargo, más recientemente, Alexeev (1967) y Wood (1993) incluyen los individuos de Olduvai en Homo habilis mientras los de otros yacimientoslos agrupos en el taxón Homo rudolfensis (del Lago Rodolfo, nombre colonial del actual Lago Turkana, en cuya cuenca se encontraron la mayor parte de los fósiles).
No hay un claro consenso sobre si Homo habilis y Homo rudolfensis representan la misma especie. Homo rudolfensis podría incluirse en H. habilis si se aceptase un amplio dimorfismo sexual (Arsuaga y Martínez, 1998). Pero Picq (2004) sugiere que la morfología del esqueleto locomotor y el tamaño relativo de los dientes no admiten diferencias morfológicas, tanto de tamaño como de forma, de tal amplitud. Los divisioniestas han propuesto que Homo habilis (sensu stricto) incluiría OH 7, 13, 16, 24, 62, KNM-ER 1501, 1502, 1805, 1813, 3735 y 42703, mientras que Homo rudolfensis incluiría KNM-ER 1470, 1590, 3732, 1801 y 1802.
Cabe destacar la clasificación que hacen Susan C. Antón, Richard Potts y Leslie C. Aiello (2014) en cuatro categorías: 1) primeros Homo, 2) grupo 1470, 3) grupo 1813 y 4) Homo habilis.
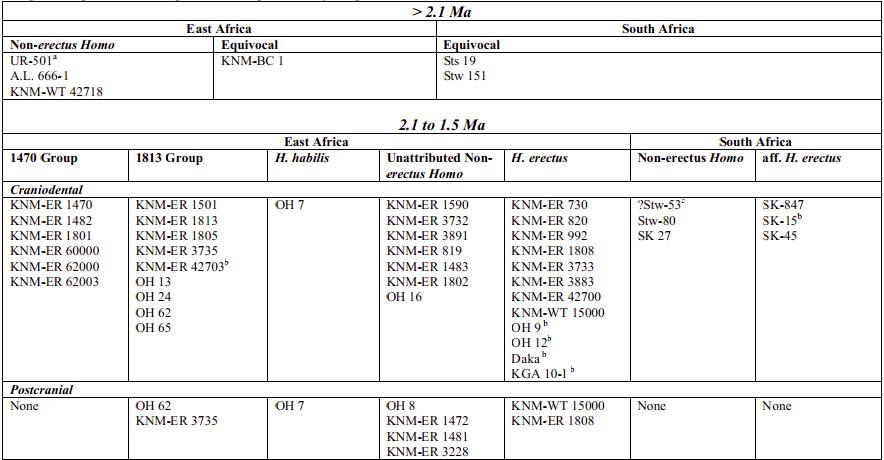
- Primeros Homo (más de 2,1 ma): respecto a Australopithecus se diferencian por la reducción en el tamaño de los dientes, la forma y tamaño de la mandíbula y la reorganización del paladar. Esta reorganización se produce de diferentes maneras: LD 350-1 (2,8-2,75 ma) muestra molares estrechos y premolares simétricos; en AL 666-1 (~2,33 ma) se reduce el tamaño de los molares, las raíces de los caninos son pequeñas, el paladar es relativamente profundo y la arcada dental es parabólica.
- ER-1470 (2,09-1,78 ma): KNNM-ER 1470 es mucho más grande que la cara de KNM-ER 62000. La forma del paladar y de la arcada los relacionan con la mandíbula KNM-ER 60000. La cara es plana y larga. Las coronas de los incisivos inferiores son estrechas con raíces relativamente cortas y la raíz canina también se reduce en tamaño en comparación con la de Australopithecus. Los premolares se reucen, pero no los molares, que son grandes. No hay reducción del tercer molar. La bóveda craneal es relativamente redondeada con un frontal verticalizado. ER 62000 tiene un tamaño del 75-80% del de ER 1470. No hay evidencias de esqueleto postcraneal.
- ER-1813 (2,09-1,44 ma): la cara de KNM-ER 1813 es anatómicamente similar a la de OH 13 y KNM-ER 1805. El esqueleto facial es bastante primitivo y relativamente prognato en comparación con 1470. La dentición anterior es algo más primitiva que en 1470, con los incisivos inferiores y raíces de los caninos más grandes. No hay reducción de la superficie oclusal del tercer molar. La bóveda craneal es redondeada en norma sagital con un frontal que muestra una constricción postorbitaria moderada. La anchura máxima del cráneo es baja, como en Australopithecus, en la región supramastoidea, pero la mayor amplitud en los parietales es sólo ligeramente inferior. Las estimaciones del tamaño del cuerpo y del encéfalo se superponen con las de ER-1470. Los restos postcraneales de OH 62 muestran una sección transversal humeral relativamente grande en relación a la femoral, lo que se ha utilizado para argumentar a favor de un componente arbóreo sustancial en el repertorio locomotor de este grupo (bipedismo facultativo como en los australopitecinos).
- Homo habilis (1,84 ma): representado únicamente por OH 7, excluido de H. erectus en base a su anatomía dental. La reconstruir la mandíbula (Fred Spoor et al, 2015) sugiriere que su morfología es incompatible con los fósiles asignados a Homo rudolfensis y con el maxilar AL 666-1. Este último es morfológicamente más derivado que OH 7, aunque es 500 mil años más antiguo. OH 7 y KNM-ER 1802 son muy similares en la forma de la arcada dental y están más cerca de OH 13 y A. afarensis que de H. sapiens y H. erectus. OH 7 conserva los dos parietales lo que ha permitido calcular una capacidad craneal de 680 cc. También conserva elementos de la mano que no tienen referencias en los otros grupos pero que permiten inferir que el individuo retiene una cantidad significativa de comportamiento arbóreo. Se diferencia en esto de H. erectus y se asemeja a ER-1813, aunque del grupo de ER-1470 no se dispone de referencias poscraneales.
Early Homo
Los materiales más antiguos atribuidos al género Homo incluyen algunos fósiles datados entre 2,8 y 2,3 ma.
LD 350-1, Ledi-Geraru, Etiopia (2,8-2,75 ma)

LF 350-1 – Ledi Geraru
El especimen LD 350-1, hallado en 2013 en Ledi-Geraru, Afar, Etiopía, es una mandíbula izquierda con dientes datada en 2,80-2,75 Ma. Combina rasgos primitivos presentes en Australopithecus con morfología derivada observada los Homo posteriores (Brian Villmoare et al, 2015). LD 350-1 está formado por dos piezas separadas por una fractura vertical al nivel del M1. Present algunas coronas dentales asociadas y fragmentos de raíz que se hallaron sueltos. Los molares son estrechos y los premolares simétricos como en los Homo posteriores. Se diferencia de Australopithecus garhi en el inferior tamaño de los dientes. El área del M2 inferior de LD 350-1 es un 64,7% de la de BOU-VP-12/130. La morfología dental y mandibular es más derivada que la de Australopithecus sediba. P4 y M3 son más pequeños que los de la mandíbula KNM-WT 8556, de asignación incierta y datada en ca 3,3 ma. LD 350-1 tiene una morfología más primitiva que H. habilis y H. rudolfensis en la porción anterior del cuerpo mandibular. LD 350-1 podría ser un miembro final de Australopithecus afarensis, con el que comparte algunas similitudes morfológicas, como las dimensiones generales de la dentición y del cuerpo mandibular y la orientación de la sínfisis mandibular, pero otras características, como la ausencia de patrones distintivos de desgaste entre el canino superior y el P3 mandibular y la profundidad uniforme del cuerpo mandibular entre el P3 y el M2, difieren de A. afarensis. La datación de LD 350-1 y la fauna asociada se ha realizado con varios métodos: datación de cenizas volcánicas mediante argón 40 / argón 39, geoquímica, paleomagnetismo y fauna. La datación ofrece ciertas dudas debido a que los fósiles se hallaron fuera de contexto. Según Erin N. DiMaggio et al (2015), la fauna asociada sugiere un hábitat abierto o mezcla de praderas mixtas y matorrales con bosques de galería bordeando ríos o humedales. El paisaje era probablemente similar al actual Serengeti o Kalahari.
Sts 19, Sterkfontein, Miembro 4, Sudáfrica (2.7-2.5 ma?)
El especimen Sts 19 consta de la base de un cráneo bien conservada hallada en los escombros de una explotación minera. Broom et al (1950) quedaron impresionados por su morfología parecida a la humana y Clarke (1977) consideró que debía atribuirse a Homo sp. en lugar de a A. africanus. Esta posibilidad fue rechazada por Dean y Wood (1982) a favor de una amplia gama de variación de A. africanus, pero Kimbel y Rak (1993) propusieron de nuevo su asignación a Homo. El hueso temporal de Sts 19, muestra una posición semejante a la humana. La forma del tímpano y los elementos petrosos y sus relaciones con la anatomía craneo-espinal no se observan en ningún otro cráneo de Sterkfontein, pero son cualitativamente similares a algunos australopitecinos robustos. La anatomía de la base del cráneo es menos derivada que la observada en los fósiles atribuidos a Homo de más de 2 ma (por ejemplo OH 13, OH 16, OH 24, KNM-ER 3891,Omo L. 894-1, STW 53), en los que la fosa mandibular se comprime antero-posteriormente y el plano preglenoideo es muy inclinado.
KNM-BC 1, Formación Chemeron, Kenia (ca. 2,4 ma)
Hill et al (1992) y Sherwood et al (2002) consideraron a este fragmento de hueso temporal, descubierto en 1966, el ejemplo más antiguo del linaje Homo basándose principalmente en la posición medial de la fosa mandibular en relación con la pared lateral de la caja craneana, que pensaban que reflejaba la expansión del cerebro, así como en varias características cualitativas de la región glenoidea y el hueso petroso. Pero se ha cuestionado la importancia filogenética de estas características ya que su presencia y expresión varían ampliamente entre los taxones de homininos fósiles y pueden ser interpretadas como simplesiomorfias. Los datos métricos de este fósil muestran afinidades tanto con Homo como con Australopithecus garhi (Asfaw et al 1999).
AL 666-1, Formación Hadar, Etiopía (ca. 2,3 ma)

AL 666-1 – Hadar
Se trata de un maxilar superior fragmentado en 35 piezas (Kimbel et al, 1996), muy probablemente de un individuo de sexo masculino, con la mayor parte de la dentición. Es similar a los fósiles de Homo habilis KNM-ER 1813, L 894-1 y a los hallazgos de Bed 1 y Bed 2 inferior de la Garganta de Olduvai en su prognatismo subnasal bajo, paladar relativamente amplio, coronas simétricas de los caninos superiores, alto ratio del área de la corona de P3/P4 superiores, M1 superior mesiodistalmente alargado y contorno oclusal romboidal del M2 superior
Carece de la morfología derivada del zigomático, zona infraorbitaria y subnasal de KNM-ER 1470 (Homo rudolfensis, Kimbel et al, 1997). Sin embargo, el P4 superior de AL 666-1 es vestibulo-lingualmente amplio en comparación con ejemplares más recientes, similar a los premolares de homininos no robustos de los niveles anteriores a 2.0 ma en la Formación Shungura, Etiopía (Suwa, 1990).
Huesos del pie, Formación Shungura (2,36-2,2 ma)
Se trata de un astrágalo (Omo 323-76-989; Miembro G; Localidad 323) y un calcáneo (Omo 33-74-896; Miembro E; Localidad 33) datados respectivamente en 2,2 y 2,36 ma (Gebo y Schwartz, 2006). El astrágalo es similar en longitud al de los humanos modernos y a KNM-ER 813, con una tróclea estrecha, un cuello corto y una cabeza estrecha tanto en términos absolutos como relativos. El calcáneo posee un gran y robusto tubérculo peroneal, es ligeramente más corto que el de los humanos modernos y es estrecho en comparación con la población de la Sima de los Huesos y los neandertales. Podrían pertenecer a Homo o a Paranthropus (Adrián Pablos, 2015).
Dientes, Miembros E-G, Formación Shungura, Etiopía (ca. 2.4-2.0 ma)

Hipótesis sobre la posición evolutiva de LD 350-1 y AL 666-1.
Suwa, White y Howell (1996) asocian estos dientes con características derivadas observadas sólo en restos de más de 2,0 ma de H. habilis y H. rudolfensis (Suwa et al, 1996). Los M1 inferiores son bucolingualmente estrechos y las coronas de los P3 inferiores son igualmente estrechas y muestran una menor asimetría y un borde marginal mesial más desarrollado que en la mayoría de los primeros australopitecinos (A. afarensis y A. africanus). Suwa et al (1996) señalaron que las coronas de los P3-M1 inferiores son relativamente grandes, con talónidos premolares expandidos, semejantes a los de KNM-ER 1802 (Wood, 1991) y UR 501 (Bromage et al 1995) (Homo rudolfensis). Se ha sugerido que este patrón dental puede haber sido primitivo para el linaje Homo.
KNM-WT 42718, Formación Nachukui, West Turkana (2,3 ma)
Homo habilis
Homo habilis surge a partir de un australopitecino grácil, sin que podamos precisar cuál. Es posible que no lo hayamos encontrado todavía. Datado en 2,5–1,6 ma. Los restos aparecieron en Olduvai a mediados de los sesenta y hasta 1964 fueron atribuidos a Australopithecus. La especie estuvo sometida a una gran polémica, que disminuyó en 1968 con el hallazgo de OH 24. En 1986 se descubre OH 62 con rasgos simiescos y las dudas volvieron a surgir. En particular, se especula sobre la posibilidad de que los restos de Olduvai pertenezcan a la misma especie que los de Koobi Fora (Homo rudolfensis) o de qué forma los restos de estos dos yacimientos se pueden asociar a especies distintas.
El holotipo de Homo habilis es OH 7 (Johnny’s child). Es un individuo joven hallado en 1960 por Jonathan Leakey, hijo de Louis y Mary. Incluye huesos parietales, mandíbula provista de 13 dientes (M3 sin erupcionar) y 21 huesos de la mano. Datado en 1,8 ma. La mandíbula está distorsionada, lo que ha obstaculizado los intentos de comparar este espécimen con otros fósiles. Fred Spoor et al (2015) reconstruyño virtualmente la mandíbula y los parietales de OH 7, y los comparó con otros fósiles. La mandíbula reconstruida muestra caracteres primitivos, con una arcada dental larga y estrecha más parecida a Australopithecus afarensis que a Homo. Esta morfología es incompatible con los fósiles asignados a Homo rudolfensis y con el maxilar AL 666-1. Este último es morfológicamente más derivado que el de OH 7, aunque es 500.000 años más antiguo. OH 7 y KNM-ER 1802 son muy parecidos y están más cerca de OH 13 y A. afarensis que de H. sapiens y H. erectus. El linaje de H. habilis se originó antes de hace 2,3 ma. De la reconstrucción de los parietales, se deduce un volumen endocraneal de 729-824 cc, mayor que el obtenido en cálculos antreriores. La de este especimen se ha estimado entre 9-13,2 años, de acuerdo con los estudios radiográficos e histológicos, la longitud de las raíces de M2 y los ratios estimados de formación de esmalte y dentina en KNM-WT 15000. La expansión del tamaño del cerebro en el Homo habilis apareció antes que los cambios en la forma de la cara.
Morfología: Capacidad craneal 610 cc (mínimo 510; máximo 824). Coeficiente de encefalización próximo a 3. Hay un aumento de la corteza cerebral, en las áreas asociativas parietales (área de Wernicke) y occipitales. El lóbulo frontal inferior (área de Broca) aumenta en superficie y complejidad. Mayor irrigación sanguínea del cerebro. Cráneo grácil con escaso desarrollo de las inserciones musculares, anchura y altura facial reducida, pilares faciales más ligeros. Huesos de la bóveda craneal delgados. Ausencia de cresta sagital. Foramen magnum en posición adelantada. Escaso prognatismo facial. La región del mentón está retrasada. Tercer molar apenas más grande que el segundo. Esmalte grueso. Un metro de estatura, peso corporal de 50 kg para los machos y 30 kg para las hembras, lo que implica un dimorfismo sexual acusado. Pudiera ser que H. habilis retuviese las extremidades superiores de un bípedo facultativo. OH 7 posee falanges proximales y medias con marcada curvatura palmar, diáfisis medio-lateral ampliada y fuertes flexores, lo que se asocia comúnmente con la locomoción arbórea. Además, la falange distal pollical de OH 7 también carece de inserción para el tendón flexor largo y otras características que se relacionan con el agarre de precisión (Moyà-Solà et al, 2008). Por tanto, las falanges de OH 7 no encajan en un patrón Homo. El pie es relativamente más corto como en los humanos modernos, con huesos parecidos a los de humanos modernos, con un hallux corto y robusto, adducido.
Lechos I y II de Olduvai (Tanzania)
- OH 4. Fragmentos del cráneo y dientes.
- OH 6. Fragmentos del cráneo y dientes.
-

OH 7 – Homo habilis
OH 7. Holotipo. Fragmentos craneales y mandíbula deformada
- OH 8. Descrito en 1960. Dos falanges, fragmento de molar y varios huesos de un pie, completo excepto el talón y la punta de los dedos, de un adulto. Su edad se estima en 1,8 ma, se desplazaba bípedamente.
- OH 13 Cinderella. Leakey, Tobias y Napier (1964). Mandíbula, dedos y fragmentos de cráneo de un adolescente, con una antigüedad de 1,6 ma. y una capacidad craneal de 650 cc.
- OH 16, George. Leakey, Tobias y Napier (1964). Huesos de los dedos y partes muy fragmentadas del cráneo. Su edad se estima en 1,7 ma. y su capacidad craneal en 640 cc.
- OH 24, Twiggy. 1968. 1,8 ma. Peter Nzube. Leakey, Clarke y Leakey. Cráneo aplastado y cinco dedos. 590 cc.
- OH 35. Fragmento distal de tibia.
- OH 62. Dik-dik hominid. 1986. 1,8 ma. Tim White. Johanson et al (1987). Incluye un paladar, fragmentos del cráneo, restos del húmero y del radio, parte del fémur y de la tibia y otros restos menores. En la zona del paladar y la dentición se parece a OH 24, ER 1813 y ER 1470. Sin embargo, las medidas de las partes superior e inferior del cuerpo son similares a las de los australopitecinos. Medía 1,05 m y sus brazos son muy largos en comparación a las piernas, con rasgos parecidos a Lucy. Adscrito a H. habilis a partir de su mandíbula y dentición.
- OH 65. 1995. Olduvai. 1,8 ma. Blumenschine et al (2003). Clarke (2012). Mandíbula superior completa y parte inferior de la cara. Datado en 1,8 ma. Por sus similitudes con ER 1470, se ha sugerido que OH 65 apoya la unificación de los dos taxones. La orientación de las estriaciones dentales de los incisivos sugiere que perteneció a un individuo diestro lo que constituye la evidencia más temprana de lateralización cerebral (Frayer et al, 2016).
- OH 86. Es una quinta falange proximal izquierda completa recientemente hallada en Philip Tobias Korongo (PTK) en la Garganta de Olduvai (Tanzania), datada en más de hace 1,84 ma, asociada a fauna y abundantes artefactos de piedra. Según Domínguez-Rodrigo et al (2015), al contrario que la falange de OH 7, OH 86 encaja en el patrón Homo. Por consiguiente, algunas morfologías de la mano primitivas persistieron al lado de morfologías parecidas a las de los humano moderno. Eeste fósil pone de relieve la coexistencia en Olduvai de Paranthropus boisei, Homo habilis y un morfo adicional representado por OH 86. Atendiendo al tamaño de OH 86, los individuos de este morfo tendrían un tamaño mucho mayor al de Homo habilis, quizá en la línea evolutiva de Homo ergaster.
- Tobias PV (2003) Encore Olduvai. Science 299(5610), 1193-1194.
- Blumenschine RJ, Stanistreet IG, Masao (2012) Olduvai Gorge and the Olduvai Landscape Paleoanthropology Project. J Hum Evol 63(2), 247-250.
Homo rudolfensis
Las características morfológicas de esta especie se parecen en unos aspectos a H. habilis y en otros a los parántropos (Wood, 2000). Presentan cerebros grandes, con capacidad media de 750 cc. El cráneo de KNM-ER 1470 muestra un ligero torus supraorbital y una constricción postorbitaria moderada. La fosa glenoidea y el conducto auditico externo están en una posición más avanzada que en Australopithecus y se observa una cara larga con la parte superior estrecha. El maxilar es cuadrado, con un paladar plano y muy corto, que presenta una gran anchura que contrasta con la de los australopitecinos. El inicio del arco zigomático está avanzado. La dentición postcanina es bastante grande, con un cociente de megadoncia de 1,5, similar a H. erectus, y las raíces son más complejas que en H. habilis, con el último molar más grande que el segundo; los grandes alveolos sugieren la presencia de potentes caninos e incisivos. Los escasos restos poscraneales disponibles sugieren un bipedismo más derivado que el de H. habilis (Picq, 2004).
Koobi Fora, Kenya (1,9 ma)
KNM-ER 1470 es el holotipo de la especie. Su capacidad craneal es de 775 cc, su bóveda craneal es más grande y menos robusta que la de los australopitecinos, no tiene cresta sagital, su cara es prominente y robusta. Una reconstrucción por ordenador realizada en 2007 determinó que el cráneo no se había reconstruido de forma adecuada y redujo su volumen a 575 cc, por lo que no tendría nada de especial y estaría dentro del rango de Homo habilis.
Cráneos
-

Cráneo de ER-1460 y mandíbula de ER 60000.
KNM-ER 1470.
- KNM-ER 1590. Wood, 1991. Incluye una parte importante de la calvaria, que se parece mucho a la de 1470 en tamaño y forma, y restos dentales de gran tamaño que, por ser dientes diferentes o en distinto grado de desarrollo, no pueden ser comparados con los de 1470.
- KNM-ER 3732. Wood, 1991. Cráneo parcial, robusto, con la parte superior de la cara y zigomático izquierdo. Los huesos frontal y parietal muestran una indicación general de mayor capacidad. En las regiones supraorbital y orbital difiere de 1470.
- KNM-ER 3891. Wood, 1991. Fragmentos craneales incluyendo fragmentos de maxilar.
Estos fósiles muestran un esqueleto facial marcadamente ortognático, con una superficie malar enorme e inclinada (Wood, 1991). KNM-ER 1470 presenta además un clivus maxilar plano por debajo de la nariz. Esta morfología facial contrasta con la de Homo habilis.
- KNM-ER 62000. Cráneo bien preservado de un joven de unos ocho años, con algunos dientes. Cara larga y aplanada. Datado entre 1,95-1,78 ma.
Mandíbulas
Los restos que se detallan son consistentes con KNM-ER 1470, pero no puede establecerse una relación directa. En algunas características, son similares a Paranthropus boisei.
- KNM-ER 819. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1482. Wood, 1991. Mandíbula.
- KNM-ER 1483. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1801. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1802. Wood, 1991. Marcado relieve en el cuerpo y eversión de la base.
- KNM-ER 60000 Mandíbula inferior casi completa de un adulto, con varios dientes y raíces. Datada entre 1,95-1,78 ma.
- KNM-ER 62003 Fragmento de mandíbula con varios incisivos. Datado entre 1,95-1,78 ma.
Estos dos últimos fósiles tienen una arcada dental corta antero-posteriormente y plana hacia delante con pequeños incisivos.
Restos poscranelaes
- KNM-ER 813. Astrágalo.
- KNM-ER 1472 (Wood, 1992). Fémur.
- KNM-ER 1473. Húmero proximal.
- KNM-ER 1481 A/B (Wood, 1992). Fémur izquierdo completo, ambas epífisis de la tibia izquierda y la inferior del peroné izquierdo, similares a los de los humanos modernos.
- KNM-ER 3228 (McHenry y Coffing, 2000). Coxis.
Otros restos
- KNM-ER 1805, The Mystery Skull. 1973. Koobi Fora, Kenya. 1,85 Ma. Paul Abell. Buena parte de un cráneo incluyendo muchos dientes. 600 cc. Con una cresta sagital parecida a la de P. boisei, pero con los dientes demasiado pequeños para pertenecer a esta especie. Podría ser asignado a casi cualquier especie pero parece más similar a Homo habilis (Wood, 1991). También se ha asignado a los australopitecinos robustos si bien diferente de cualquier especie descrita.
- KNM-ER 1813. 1973. Koobi Fora. 1,9-1,8 ma. Kamoya Kimeu. Similar a 1470, pero mucho más pequeño. 510 cc. Algunos creen que es un caso de dimorfismo sexual, otros que es una especie diferente de Homo y otros que es un australopitecino. Sorprendentemente moderno, sin cresta sagital y con prominencia nasal pequeña. Ferguson (1995) lo convirtió en el holotipo de Homo microcranous. Candidatos a paratipos serían KNM-ER 1501, 1805, 3735, 42703; OH 13, 24, 62 y 65. Para Howell (1998) y Wood (1991) pertenece a Homo habilis.
- KNM-ER 3735. 1975. Área 116, Koobi Fora. 1,9-1,88 ma. Kamoya Kimeu. Esqueleto de adulto, posiblemente macho: 14 fragmentos craneales, clavícula, escápula, radio, ulna, dos falanges de la mano, sacro, fémur y tibia. Brazos muy grandes en relación al cuerpo y potente cintura escapular, lo que parece implicar la retención de rasgos simiescos (Haeusler y McHenry, 2007). Tobias (1989), lo incluyó en Homo habilis. Para Bernard Wood (1991) se trata de Homo sp. indet.
- KNM-ER 5881. En 1980, Koobi Fora proporcionó un fragmento de fémur de Homo datado en 1,88 ma. En 2009 se recuperaron otro fragmento y un fragmento de ilion del mismo individuo.
- KNM-ER 42703. 2000. Área O, Ileret. 1,44 ma. John E. Kaatho. Maxilar derecho que incluye parte del palatino, proceso zigomático, la mayor parte del proceso alveolar y dientes de C a M3 superior. Con molares pequeños y un M3 superior corto mesio-distalmente, se separa claramente de H. habilis y H. erectus.
-
William L. Jungers et al han comunicado en la 84a Reunión Anual de la Asociación Americana de Antropólogos Físicos (2015) el hallazgo de nuevos fósiles en Kolom Odiet, Ileret, cuenca del Turkana, Kenia, durante las expediciones de 2012-2013:
-
KNM-ER 64060. Dentición inferior con todos los dientes salvo el incisivo central derecho. 2,3-2,2 ma. Se atribuye al género Homo en base a la morfología y el tamaño, posiblemente Homo habilis. Presenta los caninos más grandes registrados en los primeros Homo.
- KNM-ER 64061. Esqueleto parcial. 2,3-2,2 ma. Conserva la mayor parte de ambas diáfisis humerales, un cúbito derecho casi completo, gran parte de la clavícula derecha y la cavidad glenoidea de la escápula derecha. Los huesos largos son relativamente gráciles pero poseen corticales gruesos en sección transversal. En general, la morfología sugiere su pertenencia a un Homo temprano, posiblemente H. erectus.
- KNM-ER 64062. Esqueleto parcial. 1,86-1,82 ma. Conserva un húmero distal, un escafoides izquierdo y un pie derecho parcial que incluye un calcáneo, astrágalo, cuboides, navicular, cuneiformes, el metatarso hallucal y su falange proximal, y partes de los cuatro metatarsianos menores. Los huesos de los pies muestran una mezcla de características primitivas y derivadas.
Chiwondo Beds, Uraha, Malawi (ca. 2.5-1.9 ma)
- UR 501 es una mandíbula relativamente completa con sus dientes, de edad incierta. Tiene los premolares más pequeños que KNM-ER 1802. La inclusión en Homo se basa en la morfología de los molares y premolares y en la anatomía mandibular. Ha sido relacionada con KNM-ER 1802 (Homo rudolfensis; Bromage et al, 1995) basándose en el gran tamaño de los dientes anteriores y poscaninos, el esmalte molar relativamente grueso y las raíces dobles del P4.
- Leakey RE (1978) Koobi Fora research project. Clarendon Press, 1978.
- Walker A, Leakey RE (1978) The hominids of East Turkana. Scientific American 239(2), 54.
- Feibel CS, Brown FH, McDougall I (1989). Stratigraphic context of fossil hominids from the Omo Group deposits: Northern Turkana Basin, Kenya and Ethiopia. Am J Phys Anthropol 78(4), 595-622.
- Wood BA (1991) Koobi Fora Research Project IV: Hominid Cranial Remains from Koobi Fora. Oxford: Oxford University Press.
- Wood BA (1992) Origin and evolution of the genus Homo. Nature 355, 783-90.
- Schrenk F, et al (2002) Early hominid diversity, age and biogeography of the Malawi-Rift. Hum Evol 17(1-2), 113-122.
- Feibel CS, Lepre CJ, Quinn RL (2009) Stratigraphy, correlation, and age estimates for fossils from Area 123, Koobi Fora. J Hum Evol 57(2), 112-122.
Homo gautengensis
Sterkfontein, Sudáfrica (1,8-1,5 ma)
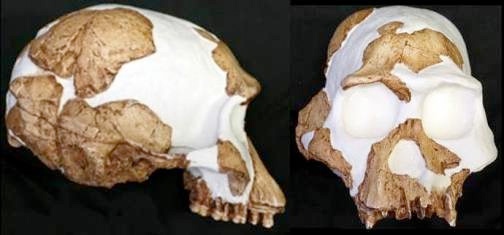
Stw-53
En 1976 Alun Hughes descubrió en la cueva de Sterkfontein un conjunto de fragmentos de cráneo y varios dientes de un individuo, Stw 53, que asignó inicialmente a Homo habilis (Hughes y Tobias, 1977). Varias décadas más tarde, Darren Curnoe, de la Universidad de Nueva Gales del Sur, con la colaboración de Phillip Tobias, clasificaron Stw 53 como el holotipo de una nueva especie: Homo gautengensis, datada en 1,8-1,5 ma (Curnoe, 2010). Su nombre proviene de la provincia sudafricana de Gauteng. Homo gautengensis se describe a partir del holotipo, descubierto por Alun Hughes, en asociación con herramientas líticas, pero se sugiere que se parece más a Australopithecus africanus que a Homo habilis (Ronald Clarke, 2013). Homo gautengensis vivió en el mismo tiempo y lugar que Australopithecus sediba, pero tiene características que descartan una posible relación. Dos nuevas revisiones de Stw 53 (Williams et al, 2012; Schroeder et al, 2014), refuerzan su inclusión en los primeros Homo, pero sin apoyar la consideración de una especie separada. Otros investigadores (Ferguson, 1989; Wolpoff, 1999) proponen su asignación a Australopithecus africanus, sobre todo por similitudes en la región medio-facial, la capacidad craneal y la región supraorbital.
- SE 255
- SE 1508
- Stw 19b/33
- Stw 75-79
- Stw 80. Mandíbula parcial aplastada y dientes. Kuman y Clarke (2000) lo asignaron a aff. H. ergaster.
- Stw 84
- Stw 151. Fragmento de un cráneo juvenil. Moggi-Cecchi, Tobias y Beynon (1998) lo asignaron a Homo temprano.
- SK 15
- SK 27. Cráneo juvenil aplastado y dientes. Howell lo asignó a Homo (1978). Curnoe y Tobias (2006) a Homo habilis.
- SK 45
- SK 847
- SKX 257/258
- SKX 267/268
- SKX 339
- SKX 610
- SKW 3114
- DNH 70
- SK 2598. Member 1 (HR). Posible P. robustus. Susman, de Ruiter y Brain (2001)
- SK 24600. Member 1 (LB). Posible P. robustus. Susman, de Ruiter y Brain (2001).
- SK 24600 está asociado al radio proximal SK 24601.
- SKX 19495, Member 3. Posible P. robustus. Susman, de Ruiter y Brain (2001).
Michael R. Lague (2015) asignó a Homo los húmeros SK 24600, SKX 19495 y SK 2598, atribuidos hasta entonces a Paranthropus robustus. El estudio no encontró base morfológica para diferenciar a Homo temprano de África del Sur de Homo habilis del Este de África, aunque los estudios de material craneodental sí lo sugieren (Grine et al, 1993; Grine et al, 2009).
Homo georgicus
En 1991 Atje Justus halló en las excavaciones de la villa medieval de Dmanisi, Georgia, la mandíbula D 211 datada en 1,8-1,7 ma, con 16 dientes, únicamente le faltan las dos ramas ascendentes. Aspecto masivo y forma estrecha, fuerte reducción del espacio retromolar y ausencia de mentón. En 1999 se hallaron: 1) La calota completa D 2280. Conserva parte de la región basal del cráneo con las partes basal y nucal del occipital ligeramente dañadas, partes del ala grande del esfenoides y la mayoría de la fosa mandibular temporal. 2) Giorgy Nioradze halló el cráneo D 2282, calota y fragmento de maxilar. Deformado. Los maxilares retienen piezas dentales ligeramente desgastadas y alveolos de dientes que son visibles en radiografía. Posiblemente perteneció a una hembra joven. En 2000 se extrajo la mandíbula D 2600. Solo le falta el extremo inferior de las ramas mandibulares. Muy grande, la mayor atribuida al género Homo; por esta razón se ha especulado con su pertenencia a otra especie diferente. Ramas mandibulares muy altas. Incisivos de tamaño muy pequeño, especialmente los centrales. Caninos grandes comparativamente. Premolares con dos raíces, lo cual es un carácter muy arcaico. Molares grandes y masivos, de esmalte grueso. Polaridad opuesta a la de D 211: tercer molar más grande que el segundo y este más que el primero. Buena parte de estas características coinciden con las de H. habilis y H. rudolfensis, pero el tamaño es mayor. Por ello, Leo Gabunia propuso la nueva especie Homo georgicus.
- Para Skinner et al (2008) las diferencias entre D 2600 por una parte, y D 211 y D 2735 por otra, son demasiado grandes para que se deban a dimorfismo sexual (Jeffrey H. Schwartz, Ian Tattersall y Zhang Chi, 2014). José María Bermúdez de Castro ha apuntado que en caso de hipotético dimorfismo sexual se daría la paradoja de que el individuo más grande, el macho, tendría una capacidad craneal mucho más pequeña que las supuestas hembras.
- Para Ann Margvelashvili et al (2013), usando las poblaciones modernas de cazadores-recolectores como referencia, las variaciones en características como la forma de la arcada dental, la altura del corpus mandibular o la inclinación de la sínfisis, permanecen dentro del rango normal. El desgaste excesivo de los dientes conduce a la ruptura de los mecanismos normales de remodelación, lo que resulta en patologías, pérdida de dientes y pérdida de la función masticatoria y además en la extraordinaria altura de la mandíbula. Sin embargo, es poco probable que ello haya limitado la duración de la vida porque este efecto fue probablemente paliado por la preparación de alimentos blandos.
- María Martinón-Torres (2013) señala la posibilidad de que esta mandíbula no sea patológica. En todo caso, las patologías habrían producido una recesión del borde superior de la mandíbula. Además resalta que del análisis del desgaste se deriva una alimentación herbívora, similar a la de chimpancés y gorilas. Esta dieta sustancialmente vegetal podría sugerir una adaptación a un nicho ecológico diferente para el individuo del cráneo 5 y los demás homínidos encontrados en el yacimiento. Además explicaría la ausencia de mecanismos compensatorios (como la migración dental mesial o mesial drift) habituales en Homo. En las mandíbulas D 211 y D 2375 se identifican estos cambios compensatorios, lo que evidencia un uso sustancialmente diferente de los dientes entre ambos conjuntos.
- Laura Martín-Francés et al (2014) detectan un desgaste dental severo e inusual, acompañado de una extensa exposición de la raíz con migración axial, abscesos periapicales y fracturas en el esmalte. Además, hay evidencia de rotación dental post-eruptiva y artropatía temporomandibular. Proponen que el patrón de desgaste observado en este individuo, similar al registrado en chimpancés y gorilas y diferente al observado en el resto de Homo, se relaciona con una dieta con un alto consumo de alimentos fibrosos y abrasivos tales como frutas y vegetales. Las superficies oclusales redondeadas y las superficies labio-linguales muy pulidas de los dientes anteriores de D 2600 podrían ser principalmente la consecuencia de actividades pre o para-masticatorias, como la sujeción y la extracción. Este tipo de comida sería también el origen de la morfología oclusal en forma de copa de la dentición posterior en combinación con un relativamente ligero desgaste proximal. Las lesiones mostradas por D 2600 no han alterado significativamente la morfología de la mandíbula y no impiden una evaluación taxonómica apropiada. Según Ann Margvelashvili et al (2016), las patologías son más prevalentes y el desgaste está más avanzado en las muestras de Dmanisi que en los actuales cazadores recolectores.
- Para José María Bermúdez de Castro et al (2014), la combinación de rasgos arcaicos y derivados en D 2600 puede estar relacionada con la dieta. Las principales características derivadas de D 2600 con respecto a las otras mandíbulas, son las siguientes:
- Presencia de un área retromolar subhorizontal y bien desarrollada.
- Alineación de corpus y ramus, con valores similares a los ejemplares de Homo ergaster (Rosas A, Bermúdez de Castro JM; 1998).
- Línea milohioidea visible e inclinada. Esta es una característica típica del linaje neandertal.
- Prominencia lateral débil al nivel de M2-M3.
- Fosa masetérica superficial.
- Borde anterior del ramus desplazado hacia delante, cubriendo solo parte de M3.
En 2001 apareció el cráneo D 2700 en extraordinario estado de conservación y la mandíbula D 2735, completa, que encaja en el cráneo anterior, que pertenecieron a un macho subadulto. Cráneo pequeño, dentro del rango de habilis. Cara baja y cóncava, de caninos prominentes y acusado prognatismo. Grandes caninos. Incisivos grandes y en forma de paleta. Caninos grandes y prominentes. En 2002 Slava Ediberidze encontró el cráneo D 3444 de 650 cc, y la mandíbula D 3900, asociados a industria olduvaiense. En 2003 se halló una mandíbula que recibió el nombre de Viejo de Dmanisi, ya que perteneció a un anciano (40-50 años) que había perdido todos sus dientes hacía mucho tiempo, por lo que debió de haber sido alimentado por otros humanos. En 2005 se halló el Cráneo 5 (D 4500; David Lordkipanidze et al, 2013; Rightmire et al, 2017) que ha resultado corresponder al mismo individuo que la mandíbula D 2600. Esta combinación mezcla de forma inaudita un cerebro pequeño (540 cc) con una cara muy prognática y exhibe afinidades morfológicas con los Homo tempranos de África. Este individuo sería un varón que medía entre 1,40 y 1,60 m y pesaba alrededor de 50 kg. Cuando murió debía tener unos 30 años. En 2011 se encontró un hueso de dedo del pie. Además se han hallado un fémur, un peroné, varias tibias, vértebras, costillas y diversos huesos de la mano y el pie. Las proporciones entre los brazos y las piernas son claramente modernas. El pie muestra un arco bien desarrollado, con el dedo gordo en posición paralela. El peso máximo sería de 50 kg y la altura máxima 1,50 m. Las palmas de las manos estaban orientadas hacia delante, lo que se interpreta como una adaptación a la vida arbórea.
Según Philip Rightmire y David Lordkipanidze (2006) los caracteres identificativos de estos fósiles son:
- Plesiomorfias (caracteres primitivos)
- Baja capacidad craneal (600-775 cc).
- Pronunciado estrechamiento frontal (índice de constricción postorbital 68,7-74,4).
- Región mastoidea inflada y proyectada lateralmente.
- Toro transversal occipital pobremente expresado o ausente.
- Índice occipital bajo: 85-102,1.
- Proyección del contorno del tercio medio facial.
- Jugum canino prominente y limitado lateralmente por el surco maxilar
- Incisura cigomáticoalveolar presente.
- Barbilla ligera o ausente.
- Sínfisis reforzada internamente (toro tansversal superior).
- Sinapomorofias con Homo erectus (senso lato).
- Toro supraorbital en forma de barra y saliente.
- Fontanela anterior con quilla sagital parietal.
- Toro angular presente.
- Escama del temporal baja, con el borde superior recto.
- Punta de la mastoides envuelta y aplanada posteriormente.
- Ángulo petrotimpánico bajo (140º-150º).
- Pirámide petrosa lisa, densa en apariencia.
- Foramen lacerum restringido.
- Silla nasal prominente.
- Posibles sinapomorfías con Homo erectus asiáticos.
- Aplanamiento parasagital (depresión) de las superficies parietales.
- Crestas paramastoidea y occipitomastoidea presentes.
- Posibles autapomorfías.
- Doble quilla sagital.
- Placa timpánica relativamente delicada.
- Proceso supratubario de Weindenreich ausente.
- M3 es de tamaño reducido.
- Gabunia L et al (2001) Dmanisi and dispersal. Evol Anthropol, Issues, News, and Reviews 10(5), 158-170.
- Vekua A, et al (2002) A new skull of early Homo from Dmanisi, Georgia. Science 297(5578), 85-89.
- Rightmire GP, Lordkipanidze D, Vekua A (2006) Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia. J Hum Evol 50(2), 115-141.
- Lordkipanidze D, et al (2007) Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 449(7160), 305-310.
- Lordkipanidze D, et al (2013) A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo. Science 342(6156), 326-331.
- Bermúdez de Castro JM, Martinón-Torres M, Sier MJ, Martín-Francés L (2014) On the Variability of the Dmanisi Mandibles. PLoS ONE 9(2), e88212.
Homo erectus / H. ergaster
KNM-ER 2598, un fragmento occipital, es el único ejemplar de H. ergaster, el taxón africano de H. erectus, más antiguo que los fósiles de Homo de Dmanisi y Java. Se ha datado en 1,87 ma. Los fósiles más antiguos de esta especie provienen de Koobi Fora (Kenia), y la especie persiste en África hasta cerca de la frontera Brunhes-Matuyama (0,78 ma). Los restos son más abundantes en Kenia (Lago Turkana) y Tanzania (Olduvai), aunque también se han hallado ejemplares en Etiopía y en Sudáfrica. Los ejemplares mejor conservados se hallaron en las riberas del lago Turkana. Tobias y von Koegniswald (1964) definieron 4 grados de hominización: 1) australopitecinos, 2) habilinos, 3) erectinos de Sangiran y ejemplares de Olduvai como OH13, y 4) erectinos de Olduvai, Ternifini, Kabuh y Zhoukoudian. De hecho, la variabilidad de las formas africanas y asiáticas se solapa en alguna medida. Dos cráneos de Turkana presentan rasgos similares a los H. erectus de Pekín (pero presentan una menor capacidad craneal): – KNM-ER 3733 (Leakey y Walker, 1985) fue descubierto por Bernard Ngeneo, del equipo de Richard Leakey, en 1975 en Koobi Fora (Kenia). Se trata de un cráneo completo de hembra madura, con capacidad craneal de 850 cc, similar a la del Hombre de Pequín. Se ha datado en ~1.63 ma (Lepra y Kent, 2015); – KNM-ER 3883 (Leakey y Walker, 1985). Cráneo de 800 cc datado en 1,6 ma. Si bien muchos paleoantropólogos diferencian entre H. ergaster y H. erectus, sus similitudes y la dificultad de asignar algunos restos a uno de los dos taxones recomiendan considerar estas especies en conjunto formando lo que podemos llamar el grado erectus.
- Homo erectus, Dubois, 1892; Weidenreich, 1940. Incluye los hallazgos de Dubois en Java a finales del XIX y los de China en la década de 1920. Weindenreich estableció los criterios diagnósticos en las décadas de 1930 y 1940. En 1950, Ernst Mayr propuso agrupar la mayoría de los fósiles atribuidos al género Homo conocidos hasta entonces en Homo erectus.
- Homo ergaster, Groves y Mazák, 1975. Holotipo: KNM-ER 992 (mandíbula). Homo ergaster fue propuesto en 1975 por Groves y Mazák y su uso se generalizó gracias a los trabajos de Wood en los 1980.
A medida que los nuevos hallazgos se han ido acumulando, para los taxónomos agrupacionistas (lumpers) el grado erectus sensu lato ha ido reuniendo ejemplares tan diferentes como el Homo georgicus o Homo floresiensis. Emiliano Bruner et al (2014) analizó la variación endocraneal de especímenes africanos y asiáticos, sugiriendo que los ejemplares de China, Indonesia y África se superponen en gran medida en su distribución fenotípica y en sus patrones morfológicos. Igulamente, Baab (2016) concluyó que los H. erectus de China, Indonesia, Georgia y el este de África comparten una forma neurocraneal distinta de la de otros taxones del Plio-Pleistoceno. En la variación morfológica del grado erectus, los ejemplares de Indonesia forman una puente entre las poblaciones anteriores y posteriores de África y Asia. En cualquier caso, KNM-ER 2598, un fragmento occipital africano, es anterior a los fósiles de Dmanisi y Java, lo que indica que se originaron en África del Este hace 1,9 ma o antes y después emigraron al continente Euroasiático. El registro fósil sugiere que los hominini de cerebro grande (más de 800 cc) aparecen hace 1,8 ma, coincidiendo con la acentuación de la aridez y la pérdida de vegetación que dan inicio al Pleistoceno. La respuesta a esta presión ambiental fue un aumento alómetrico del tamaño corporal, del volumen del cerebro y reducción del dimorfismo sexual. Fuertes afinidades craneofaciales sugieren que AL 666 podría haber formado parte de la población de origen de este taxón (Susan C. Antón, 2012). El cerebro sigue creciendo lentamente hasta alcanzar 1.100 cc en los ejemplares más recientes. Junto con el aumento de cerebro, se incrementan también las capacidades cognitivas: al principio H. erectus mantiene la tecnología olduvaiense (modo 1) de los H. habilis. hasta que aparece una nueva tradición cultural: la achelense (modo 2), ausente en Asia, con alguna excepción, y que no aparece en Europa hasta época tardía (0,5 ma). Homo erectus migró rápidamente a Asia (1,8-1,7 ma). En el sudeste de Asia persistió hasta el Pleistoceno Final. El grado persiste en África durante más de un millón de años, hasta hace 0,78 ma.
Danakil, Afar, Etiopía
El norte del Danakil (Afar, Eritrea) ha proporcionado evidencia de H. erectus en la cuenca del río Dandiero, al sur de Buia (Abbate et al, 1998). Las excavaciones corren a cargo de la Expedición Danakil Italiana y Eritrea, coordinada por la Universidad La Sapienza de Roma y el Museo Nacional de Eritrea, bajo la dirección de Alfredo Coppa, dentro del Proyecto Internacional Buia.
El yacimiento de Uadi Aalad (UA), consta de depósitos deltaicos y fluviales de 5-6 m de espesor radiométricamente y biocronológicamente datados en ca. 1 ma (Ghinassi et al, 2009). Destacan los hallazgos en la capa LK3 Homo (Macchiarelli et al 2004; Bondioli et al, 2006).
- UA 31. Buia. Cráneo femenino adulto prácticamente completo, con la cara, con mezcla de características erectus/ergaster y rasgos derivados, incluyendo caja craneal larga y ovalada, de más de 20 cm de longitud. Según Bruner et al (2016), su capacidad craneal es de 995 cc.
- UA 222 y UA 369. Dientes incisivos permanentes. Tienen un esmalte relativamente delgado en comparación con la condición humana moderna, como en Homo antecessor, y la morfología recuerda al H. heidelbergensis norteafricano.
- UA 173, UA 405 y UA 466. Fragmentos de pelvis.
En Mulhuli-Amo (MA), a 4,7 Km de Uadi Aalad y formando parte posiblemente del mismo horizonte estratigráfico se han hallado nueve fragmentos humanos, pertenecientes posiblemente a tres individuos.
- MA 14 fragmento de frontal.
- MA 64 y MA 88 a-f, fragmentos parietales y MA 89 fragmento temporal, pertenecientes al mismo individuo adulto. MA-88, compuesto por los seis fragmentos a-f, constituye un parietal izquierdo casi completo cuyo espesor y características estructurales se parecen a las de UA 31.
- MA 93 Corona sin uso de un molar permanente inferior.
La campaña de 2016 ha proporcionado huellas de erectus datadas en hace 800 ka, en los sedimentos fluviales de un paleolago rodeado de pastizales, en el yacimiento de Aalad-Amo.
Daka (Dakanihylo), Formación Bouri, Middle Awash, Etiopía
- BOU-VP-2/66. Asfaw et al, 2002. Caja craneana descubierta por Henry Gilbert, del equipo de Berhane Asfaw y Tim White, en 1997, datada en 1 ma y con una forma similar a la de Buia. Según Carlson et al (2013) es similar en morfología lineal a los fósiles de H. erectus asiáticos. Para Baab (2016), pudo haber pertenecido a una población «avanzada» de H. erectus cercana a la raíz del Homo heidelbergensis sensu lato, o a una población inicial de H. heidelbergensis sensu lato.
Olorgesailie
- KNM-OG 45500. Cráneo parcial datado en 0,9 ma, descubierto en 2003 por el equipo de Rick Potts. Capacidad craneal de unos 700-800 cc. Se recuperó aproximadamente 1,5 Km al este de una gran acumulación de hachas de mano, que provienen de la misma capa.
Gombore I (Melka Kunture), Etiopía
- MK3. En las excavaciones dirigidas por J. Chavaillon en 1976 en Gombore I (Melka Kunture, Etiopía) apareció in situ, en la unidad 2 del nivel 3, datada en >1,39 ma, una porción distal bien conservada de un húmero izquierdo de hominini conocido como Gombore IB-7594, Melka Kunture 3 o MK3, asociado a un conjunto lítico de transición olduvaiense a achelense. Fabio Di Vincenzo et al (2015) indican que MK3 añade una gran cantidad de variabilidad al género Homo. El gran tamaño de MK3 sugiere un peso corporal cercano a 90 Kg, lejos de la gama del tamaño del cuerpo conocido para los Homo del Pleistoceno temprano.
Olduvai, Tanzania (1-0,6 ma)
- OH 9, Chellean Man (Heberer, 1963; Rightmire, 1979). Descubierto por Louis Leakey en 1960 en la garganta de Olduvai (Tanzania). Su edad se estima en 1,5 ma. Consiste en una bóveda craneal de unos 1065 cc de capacidad.
- OH 12, Pinhead (Rightmire, 1979). Descubierto por Margaret Cropper en 1962 en la garganta de Olduvai (Tanzania) Es parecido, pero menos completo que OH 9, y su capacidad craneal es menor, 750 cc. Muy similar a KNM-ER 3733, aunque un millón de años más joven. Su edad está comprendida entre 600 y 800 Ka.
- OH 23, fragmento de mandíbula, datado en 0,6 ma.
- OH 28 (Rightmire, 1990). Coxis.
- OH 34 (Rightmire, 1990). Fémur.
- OH 82 (Hlusko, Reiner y Njau, 2015). Ulna. Sedimentos datados en hace ca 1 ma.
Turkana, Kenia
- KNM-ER 730 (Wood, 1991). Occipital, parietal, frontal y mandíbula parciales.
- KNM-ER 736 (Rightmire, 1990). Fémur.
- KNM-ER 737 (Rightmire, 1990). Fémur.
- KNM-ER 820 (Wood, 1991). Mandíbula de subadulto.
- KNM-ER 992 (Wood, 1991). Mandíbula. Holotipo de Homo ergaster.
- KNM-ER 1472. (Wood y Collard, 1999). Fémur.
- KNM-ER 1481. (Wood y Collard, 1999). Fémur.
- KNM-ER 1808 (Walker, Zimmereman y Leakey, 1982). Esqueleto y fragmentos craneales.
- KNM-ER 3228. Coxal.
- KNM-ER 3733 (Leakey y Walker, 1985). Cráneo.
- KNM-ER 3883 (Leakey y Walker, 1985). Cráneo.
- KNM-ER 42700 (Spoor et al, 2007). Calota craneal de adulto joven hallada en Ileret, Kenia, datada en 1,55 ma. Su atribución taxonómica está sujeta a controversia (Baab, 2016). El tamaño es de los más pequeños conocidos para H. erectus (Catherine C. Bauer y Katerina Harvati, 2015).
- KNM-WT 15000, Turkana Boy Descubierto por Kamoya Kimeu en 1984 en la ribera del río Nariokotome cerca del lago Turkana (Kenia) (Brown et al. 1985; Leakey and Lewin 1992; Walker and Leakey 1993; Walker and Shipman 1996). Se trata de un esqueleto casi completo de un niño entre 7,6-8,8 años (Dean y Smith, 2009) al que le faltan las manos y pies. Es el Homo erectus más completo conocido, con una edad de 1,5 ma. Su capacidad craneal es de 880 cc, y hubiera alcanzado los 910 cc de completar su desarrollo. Su talla era de 1,60 m. Christopher B. Ruff y M. Loring Burgess (2014) han estimado una estatura adulta de 176-180 cm y una masa corporal adulta de 80-83 Kg en base a un patrón de crecimiento similar al de los simios africanos. Los músculos debían de estar muy desarrollados. Excepto por el cráneo. La forma y el tamaño del canal espinal son similares a los de los humanos modernos (Meyer y Haeusler, 2015).
- KNM-WT 51260, tercer metacarpiano hallado en Kaito (oeste del Lago Turkana) en 2012, datado en 1,4 ma. Según el estudio de Carol WARD V. et al (2013) el hueso se asemeja al de un humano moderno en proporciones generales y morfología.
Otros materiales de África
- BSN49/P27. Gona (Simpson et al, 2008). Pelvis. Debió pertenecer a una hembra de tamaño pequeño, pero con una cadera amplia para ese tamaño.
- SK 15 (Robinson, 1961; Howell, 1978; Grine, 2001). Mandíbula parcial.
- SK 45 (Robinson, 1961; Howell, 1978; Grine, 2001). Mandíbula parcial.
- SK 847 (Clarke, Howell y Brain, 1970; Tobias, 1991; Walker, 1981; Kimbel, Johanson y Rak, 1997; Grine, 2001). Restos mandibulares, dentales y cráneo parcial.
- SK 1896 (Susman, de Ruiter y Brain, 2001). Fragmento distal de fémur.
- SK 2045 (Susman, de Ruiter y Brain, 2001). Fragmento proximal de radio.
- SKW(SKX) 34805 (Susman, de Ruiter y Brain, 2001). Fragmento distal grande de húmero.
- GAR IVE (Condemi, 2004; Zanolli et al, 2016). Mandíbula parcial inmadura hallada en Garba, Melka Kunture, en 1981. Se trata del representante humano más antiguo en un entorno montañoso, datado en hace ca 1,7 Ma.
Out of Africa I
Los homininos del grado erectus no son los primeros humanos en salir de África. Homo georgicus le precedió. La evidencia de la Cueva Longgupo (donde se halló un fragmento mandibular con un premolar de rasgos primitivos) sugiere que los primeros hominini en ocupar Asia fueron cercanos a H. habilis por su reducido tamaño corporal (Manuel Will y Jay T. Stock, 2015). Las dataciones de Han Fei et al (2015) han proporcionado fechas de ca 1,35 ma. María Martinón-Torres (2006) propone que H. erectus en Ásia se origina a partir de una población asiática relacionada con estas primeras expansiones. En Quranwala, India, se hallaron herramientas líticas y huesos con marcas de corte datados en 2,8 ma (Dambricourt, 2016). En Yiron (Israel) se han hallado herramientas olduvaienses, datadas en 2,4 ma, pero el conjunto es muy discutido. Las industrias líticas de Soan Valley, Riwat, Pakistán, se han datado en 1,9 ma. Erq el-Ahmar (Israel, Ron y Levi, 2001), con herramientas olduvaienses, está datado entre 1,9-1,7 ma. Estas tecnologías pudieron tener procedencia africana, pero no se ha descartado un origen asiático independiente. Los hallazgos de Mojokerto y Sangiran (Java) se han datado en 1,8-1,6 ma, pero están sujetos a fuerte controversia. En Yuanmou, sur de China, se hallaron dos incisivos centrales, presumiblemente de H. erectus y artefactos líticos datados en 1,7 ma. En la cuenca de Nihewan, al noreste de China, hay indústrias líticas datadas en más de 1,5 ma:
- Majuangou (MJG). Se hallaron también huesos de caballo con marcas de percusión, rotos para la extracción del tuétano. ~ 1.70 ma para MJG-III y MJG-II; ~ 1.40 ma para MJG-I.
- Shangshazui (SSZ). 1,7-1,6 ma.
- Lanpo (LP). 1,6 ma.
- Xiaochangliang (XCL). 1,36 ma.
- Xiantai (XT). 1,36 ma.
- Banshan (BS): BS ~ 1.35 ma.
- Feiliang (FL). 1,2 ma.
- Donggutuo (DGT). 1,1 ma.
- Ubeidiya (Israel). Datado en 1,4 ma, con grandes hachas de mano (González y Moure, 2000; Goren-Inbar et al, 2000).
- Pabbi Hills, Paquistán, cuenta con herramientas líticas datadas en 1,4-1,2 ma.
- Kocabaş (Turquía), cráneo con una combinación de rangos arcaicos y evolucionados a medio camino entre los fósiles de Dmanisi y los de Zhoukoudian. Por su morfología y dimensiones, está cerca del H. erectus de África. Se diferencia del H. erectus de China por las menores proporciones de su escama del frontal y una constricción post-orbitaria marcada. Se ha datado en 1,3-1,1 ma (Anne-Elisabeth Lebatard, 2014). Sedimentos del río Gediz sugieren una ocupación entre ca 1,24-1,17 ma (Maddy et al, 2015).
- Kuldara (Khovaling, Sur de Tayikistán) fue excavado entre 1981-1984 y ha ofrecido 96 instrumentos líticos y algunos artefactos de hueso. Está datado en 0,85 ma (Vadim A. Ranov, Eudald Carbonell y Xosé Pedro Rodríguez, 1995).
- El yacimiento de Karama (Altai) está datado en 1-0,8 Ma.
Esta cronología coloca a China en la misma cronología que Java y Georgia e indica una rápida propagación de H. erectus por Asia. Los yacimientos en el norte y centro de Asia muestran ocupaciones intermitentes dependientes del ciclo glaciar. El modo tecnológico 1 persiste de forma muy dilatada en el tiempo y coexiste con el modo 2 cuando este aparece tardíamente. En el sur de Asia el modo 2 se desarrolló más temprano. Homo erectus pudo penetrar en Asia por el Corredor de Palestina, siguiendo la misma ruta que utilizó el grupo que dio lugar al Homo georgicus, pero también pudieron hacerlo atravesando el istmo de Bab-el-Mandab, próximo a Hadar (Etiopía). De acuerdo con el modelo desarrollado por Carotenuto et al, 2016, H. erectus siguió casi de forma pasiva a los grandes herbívoros durante su dispersión. Durante la fase inicial de la dispersión, evitó las zonas densamente pobladas por grandes carnívoros y prefirió las zonas de latitud baja o media más cálidas. Durante la fase posterior, asociada a la indñustria achelense, H. erectus se asoció con piedra caliza y margas arcillosas, y parece haber seleccionado lugares elevados.
China – 1,8-1,2 ma
- Yuanmou, Danawu
-
La capa 25 ha proporcionado dos incisivos (Hu, 1973) datados por paleomagnetismo en 1,7 ma (Zhu et al, 2008).
- Una tibia izquierda fue recogida en la ladera sur de Guojiabao (Zhou, 1998), en un estrato que puede ser equivalente a la capa 27.
-
- Tiandong, Mohui, Guangxi Zhuang
- Segundo molar inferior derecho completo en la capa 3.
- Jianshi, Longgudong, Hubei
- Tres dientes: tercer molar superior izquierdo (capa 8 de la sección A-A’), tercer premolar superior derecho (capa 5 de la sección G-G’) y primer molar inferior derecho (capa 8 de la sección C-C’). Datación de 2,14 Ma (Gao y Cheng, 2004).
- Lantian, Gongwangling, Jiujianfang
-
En 1964, un equipo del Instituto de Paleontología de Vertebrados y Paleoantropología de Pekín halló en una pequeña colina de Gongwangling (Lantian, Shaanxi) restos de Homo, semejantes a los erectus más antiguos de Java (Wu y Poirier, 1995; Wu et al, 1966). Apareció primero un molar superior aislado y varios meses más tarde grandes fragmentos de la bóveda craneal. También se recuperó una gran cantidad de material de fauna de mamíferos. Un predominio de especies tropicales y subtropicales ha sugerido un clima más cálido que hoy (Gu y Jablonski, 1989; Qi, 1989). Se han hallado artefactos líticos (Dai, 1966; Tai y Hsu, 1973). Los materiales óseos de Homo fueron descritos por Woo (1965) y Wu y Poirier, 1995. Los fósiles humanos (PA 1051-6) incluyen un frontal completo, gran parte de los parietales, la mayor parte del temporal derecho, parte de los nasales, una gran parte del maxilar derecho con segundos y terceros molares asociados, y parte del maxilar izquierdo. La preservación de los fragmentos de hueso es extremadamente pobre. Superestructuras muy marcadas: torus frontal y occipital, gran espesor de los gruesos del cráneo, constricción postorbitaria marcada, boveda craneal alargada y baja. 770 cc.
-
Zhao-Yu Zhu et al (2014) han datado los fósiles en ca 1,62 ma, probablemente ca. 1,63 ma, lo que hace de Gongwangling el segundo sitio más antiguo fuera de África con restos craneales (después de Dmanisi).
-
China – 1,3-0,8 ma
- Yunxian, Quyuanhekou, Hubei
- Hallazgos en la capa 3, datados en 1,1-0,9 ma (Vialet et al, 2011): 1989, Yunxian I. Cráneo. 1990, Yunxian II. Craneo de 1.100 cc asociado a restos de fauna e industria lítica. Ambos cráneos están muy deformados, son muy robustos y tienen un perfil bajo y alargado. Dennis A. Etler ha destacado las similitudes con ATD6-69 (Atapuerca, Gran Dolina).
China – 0,850-0,500 ma
- Zhoukoudian (inferior), Fangshan, Beijing
- En la cueva Peking Man Site de Zhoukoudian (Pequín), excavada por J.G. Andersson, entre 1929-1937 se encontraron 5 bóvedas craneales, 14 mandíbulas fragmentarias, 147 dientes aislados y huesos de esqueletos, que fueron inicialmente atribuidos al Sinanthropus pekinensis. La cueva fue ocupada por Homo y hienas de forma intermitente.
- Los restos más antiguos (capas 8-11) podrían llegar hasta hace 750 Ka (Shen et al, 2009). Las capas 3-7 podrían tener más de 400 Ka (Shen et al, 2001) si bien los análisis bioestratigráficos proporcionan dataciones inferiores (Wei Dong, 2016).
- Los restos más completos corresponden a Skull III, descubierto en Locus E en 1929, es el cráneo de un adolescente con capacidad craneana de 915 cc; Skull II, descubierto en Locus D en 1929, es un adulto o adolescente con capacidad de 1030 cc; Skulls X, XI and XII (antes llamados LI, LII y LIII) fueron descubiertos en Locus L en 1936, se trata de un adulto masculino, una hembra y un jóven, con capacidades de 1225 cc, 1015 cc y 1030 cc respectivamente. (Weidenreich 1937).
- Los fósiles los empezó a estudiar Davidson Black hasta su muerte en 1934. Franz Weidenreich lo reemplazó. Los fósiles desaparecieron en 1941 durante la II Guerra Mundial al ser embarcados, por seguridad, hacia USA, pero quedan algunos moldes y las descripciones de los autores anteriores.
- Skull V: dos cráneos descubiertos en 1966 y otros fragmentos descubiertos en 1934 y 1936 de bóvedas craneanas con una capacidad de 1140 cc. (Jia and Huang 1990).
- En 2015 se comunicó la identificación de un canino de adulto hembra desgastado y rasgado procedente de este yacimiento en una caja en el Museo de Uppsala.
- Yunxian, Meipu
- Tres dientes.
- Luonan, Longyanandong, Donghe, Chengguang, Shaanxi
- Primer molar superior derecho. Xue, 1987.
- En la cueva paralela (Longyabeidong), aparecieron gran cantidad de artefactos líticos. Se considera que la fauna asociada a ambos hallazgos es equivalente.
- Yunxi, Bailongdong, Shenwuling, Anjia, Hubei
- Ocho dientes en la capa 2 (Wu et al, 2009; Liu et al, 2014).
China – 0,600-0,300 ma
- Longtandong, Hexian, Anhui, Este de China. Capa 2
- 1980-1982. Calota casi completa, dos fragmentos craneales, corpus mandibular izquierdo parcial con M2 y M3, diez dientes aislados y restos poscraneales fragmentarios de cuatro individuos, datados en 300 Ka (Huang et al, 1995).
- Hexian I: neurocráneo hallado en 1982 (Cui y Wu, 2015).
- PA-831. Fragmento de mandíbula (Wu Liu et al, 2017).
- Los dientes son primitivos métrica y morfológicamente y se superponen con los de H. ergaster del Este de África en sus grandes dimensiones y complejidades oclusales (Song Xing et al, 2014).
- Chenjiawo, Xiehu, Lantian, Shaanxi
- Lantian 1, una mandíbula carente de las últimas muelas, datada por estratigrafía en 0,65 Ma (An y Ho, 1989). Capa 5. Zhang et al, 1964.
- Yiyuan, Qizianshan
- Fragmento craneal y cinco dientes en la Localidad 1 y otros cinco en la Localidad 2. 0,4-0,3 ma. El estudio comparativo métrico y morfológico realizado por Song Xing et al (2016) relaciona los dientes de Yiyuan con las muestras de Zhoukoudian, Chaoxian y Hexian. Estas muestras son menos derivadas que las de Panxian Dadong (H. sapiens arcaico).
China – 0,400-0,150 ma
- Nanzhao, Xinghuashan, Ruanzhuang, Yunyang, Henan
- Cuarto premolar inferior derecho. Capa 5.
- Huludong, Nanjing, Leigong, Tanshan, Jiangning, Jiansu
- 1993. Nanjing 1 (Capa 2) y Nanjing 2 (Capa 5). Dos cráneos fragmentarios.
- Datados en 500 Ka (Wang et al, 2002).
- Dongzhi, Hualongdong, Wang, Raodu, Anhui
- Dos fragmentos craneales y dos dientes asociados a artefactos. Capa 3. Gong et al (2014).
- Chaoxian. Un occipital y un maxilar con seis dientes datados en 0,2-0,16 Ma.
- Penghu 1 es un fragmento mandibular derecho con los premolares y molares muy desgastados. Datado entre 190-130 Ka o 70-10 Ka. Estas fechas son inesperadamente tardías para la apariencia robusta y primitiva de Penghu 1, desconocida en el registro asiático con excepción de Hexian (Chun-Hsiang Chang et al, 2015).
Poblamiento de Asia
José María Bermúdez de Castro defiende diferentes eventos de colonización de Asia. En base al estudio de los dientes de Hexian (Song Xing et al, 2014) sugieren escenarios evolutivos complejos de diferentes linajes en Eurasia. Hexian podría representar la persistencia en el tiempo de un H. erectus que habría conservado rasgos primitivos que se perdieron en otras poblaciones asiáticas como Zhoukoudian o Panxian Dadong (Wu Liu et al, 2017). La inestabilidad climática y el cambio ambiental en Asia podrían haber favorecido un patrón de fragmentación y aislamiento con alta variabilidad morfológica.
Los dientes de Hexian son primitivos métrica y morfológicamente y se superponen con los de H. ergaster. Zhoukoudian es morfológicamente más derivado, pese a ser contemporáneo o ligeramente más antiguo (Shen et al, 2009). Hexian podría ser también más primitivo que Chaoxian que habría retenido características primitivas perdidas en otras poblaciones contemporáneas. El hecho de que los fósiles asiáticos no compartan con H. antecessor y el linaje neandertal algunas características dentales apunta a una divergencia temprana entre las poblaciones europeas y asiáticas del Pleistoceno temprano.
Chun-Hsiang Chang et al (2015) proponen varios modelos para explicar la persistencia de Penghu 1, con una morfología tan arcaica, hace menos de 190 Ka.
- La retención de características de los Homo tempranos de Asia debido a que los H. erectus de Java y China muestran un corpus mandibular más delgado y molares más pequeños, esta hipótesis implica la presencia de otro linaje en Asia que se prolongó desde el Pleistoceno Temprano.
- Una migración de un grupo africano con mandíbula robusta, posiblemente trayendo consigo la tecnología lítica achelense, alrededor del final del Pleistoceno Inferior, que más tarde evolucionó a nivel local. Esto es compatible con la sugerencia reciente de que el Homo arcaico tardío chino recibió alguna influencia genética del oeste y puede ser incluido en H. heidelbelgensis. Penghu 1 parece ser más primitivo y similar a Hexian cuya morfología craneal es distinta de Jinniushan y otros Homo arcaicos tardíos chinos.
Ambas hipótesis ponen en duda la idea tradicional de que H. erectus fue la única especie de hominini en el continente asiático en el Pleistoceno Inferior y principios del Pleistoceno Medio, que exhibió variación geográfica clinal en todo el continente desde Indonesia hasta el norte de China.
Otros hallazgos fósiles en China han descubierto fósiles más modernos que apoyan la coexistencia en esa zona de Homo erectus y Homo sapiens, o al menos la evolución asiática de estos taxones. Para Howell constituyen un paleodemo diferenciado. Restos que se asimilan por su parecido y capacidad craneal a H. sapiens se han datado en unos 0,58 ma, lo que supondría que el H. sapiens asiático ha evolucionado antes que los H. sapiens europeos.
- Dali. 1978. Cráneo completo datado en 0,3-0,25 ma por U-sereis. Parietal más verticalizado, con fosas postcanina y maxilar pronunciadas, concavidad en el pómul y rostro corto y vertical, muy poco prognato. Con rasgos parecidos a los restos hallados en Zhoukoudian pero también con rasgos modernos. 1.150 cc.
- Miahoushan. Restos datados entre 0,22 y 0,14 Ma.
- Changyang
- Localidad 4 de Zhoukoudian
- Yanghuidong (Tongzi, Guizhou). Dientes y fragmentos craneales. 0,2 ma.
- Hathnora (Narmada, India). Cráneo datado en 0,2 ma. Presenta un mosaico de caracteres de H. erectus y H. sapiens. Bóveda craneal más elevada y superestructuras menos marcadas.
- Dingcun. 0,11 ma.
- Hualongdong. (Jiandé, Dongzhi, Anhui). 0,09 ma. El 11 de octubre de 2015 apareció un cráneo (Dongzhi Man), junto con una variedad de instrumentos de piedra, dientes y fragmentos de hueso humano de al menos cuatro individuos, uno de ellos infantil, así como más de 6.000 fósiles de vertebrados. Se han datado entre 0,412-0,150 ma.
- Guojiujan.
- En la Región de las Tres Gargantas se han localizado 10 yacimientos de hominini y otros 30 líticos (Modo 1) y con otras evidencias de actividades humanas. Yacimientos más importantes: Yandunbao, Miaogou, Chibaling, Ranjialukou, Gaojiazhen, Fanjiahe, Wuyang, Cueva Xinglong, Cueva Sunjia, Cueva Zhongjiawan, Jingshuiwan, Zaoziping, Dadiping, Outang, Cueva Leiping, Cueva Migong y Cueva Huanglong (2004-2006, 7 dientes asociados a herramientas de piedra y hueso y a fósiles de mamíferos, datados en 0,1 ma).
Howell distingue un paleodemo adicional para Jinniusham (Liao Ning, Manchuria). Esqueleto femenino incompleto, único hallado en Asia correspondiente al Pleistoceno Medio. 1984. Datado por ESR en 0,28 ma. Gran anchura de la pelvis. Pudo pertenecer a un individuo de 168 cm y 78 Kg. El cráneo, de 1.330 cc, conserva rasgos primitivos, como la bóveda craneana todavía baja y alargada hacia la parte posterior.
- Weidenreich F (1943) The skull of Sinanthropus pekinensis: a comparative study on a primitive hominid skull.
- Wang YY, et al (1979) Discovery of the fossil Dali Man in Shaanxi Province and the preliminary study. Chinese Science Bulletin 24, 303-306.
- Wu XZ (1981) A well-preserved cranium of an archaic type of early Homo sapiens from Dali, China. Scientia Sinica 24(4), 530.
- Pope GG (1992) Craniofacial evidence for the origin of modern humans in China.» Am J Phys Anthropol 35(S15), 243-298.
- Etler DA (1996) The fossil evidence for human evolution in Asia. Annual Rev Anthropol 275-301.
- Weiner S, et al (1998) Evidence for the use of fire at Zhoukoudian, China. Science 281(5374), 251-253.
- Goldberg P, et al (2001) Site formation processes at Zhoukoudian, China. J Hum Evol 41(5), 483-530.
- Shen G, et al (2009) Age of Zhoukoudian Homo erectus determined with 26Al/10Be burial dating. Nature 458(7235), 198-200.
- Wu X, Sheela A (2013) A description of the geological context, discrete traits, and linear morphometrics of the Middle Pleistocene hominin from Dali, Shaanxi Province, China. Am J Phys Anthropol 150(1), 141-157.
Java
Dubois descubrió cerca de Trinil una calota en 1891 y un fémur en 1892, a partir de los cuales definió el taxón Pithecanthropus erectus. Swisher et al (1994) dataron los restos de Mojokerto en 1,81 ma y los de Sangiran en 1,6-1,1 ma. En 2011, Russell L. Ciochon, Yahdi Zaim y E. Arthur Bettis dataron en 1,5 ma unos restos de mandíbula hallados en Sangiran. Josephine C. A. Joordens et al (2014) dataron sedimentos con conchas de mejillones de agua dulce de Trinil mediante 40Ar/39Ar y luminiscencia, obteniendo una edad máxima de 0,54 ± 0,10 ma y una edad mínima de 0,43 ± 0,05 ma, lo que supone una fecha mucho más joven que la considerada hasta ahora. La presencia de estos Homo en Java plantea el problema de cómo cruzaron los brazos de mar entre las islas:
- Hipótesis de la desecación de ciertos mares en esa época. Java está unida al continente por una plataforma continental poco profunda.
- Se han encontrado elementos líticos y al Homo floresiensis en la isla de Flores que se encuentra aislada por aguas muy profundas. Si Homo erectus fue capaz de construir balsas hay que concederle una capacidad cognitiva muy elevada. Los restos de Flores se han datado en 75 Ka.
Trinil
- Java Man, Pithecanthropus I, (Pithecanthropus erectus). Descubierto por Eugene Dubois en 1891 cerca de Trinil (Java). Holotipo de Homo erecuts. Su edad es de menos de 1 ma. Calota gruesa con un perfil muy bajo, frontal muy huidizo y torus supraorbitario relativamente poco marcado. Capacidad craneana 940 cc. Atribuido al sexo femenino.
- Trinil 3 (Fémur I). Fémur izquierdo completo pero patológico hallado en agosto de 1892. Para Ruff et al (2013, 2015) no corresponde con H. erectus y es más joven que el resto de la colección. Los niveles en el hueso de Ca y P indican que fue extraído de un nivel estratigráfico diferente (Bartsiokas y Day, 1993).
- Posteriormente se recuperaron otros cuatro fémures (Trinil Fémures II-V): Trinil 6 (Fémur II), Trinil 7 (Fémur III), Trinil 8 (Fémur IV), Trinil 9 (Fémur V).
Ngandong
- Solo: investigadores holandeses descubrieron más fósiles en la década de 1930 cerca de la aldea de Ngandong, en el río Solo. Hasta hace poco, se pensaba que representaban una población muy reciente, pero el trabajo de campo y los análisis más recientes sugieren que aquellos Homo vivieron en algún momento entre 143-546 Ka. 11 calotas y 2 tibias. Los cráneos tienen las superestructuras muy marcadas y sus bóvedas son ligeramente altas. Capacidades en torno a 1.150 cc. La morfología parece corresponder a un estadio evolutivo final del H. erectus. Datados en 0,05 Ma.
- Ngandong-6. 1.251 cc.
- Ngandong-7. 1.013 cc.
- Ngandong-10. 1.135 cc.
- Ngandong-11. 1.231 cc.
- Ngandong-12. 1.090 cc.
- En Ngandong y en la localidad próxima de Matar se hallaron lascas y algunos núcleos rudimentarios (Fauzi et al, 2016).
Mojokerto
- El niño de Mojokerto es un cráneo infantil descubierto en Perning (Mojokerto) en 1936, de 630 cc. Torus supraorbitario incipiente. La estiomación de la edad de muerte varía desde 1 hasta 8 años.
Sangiran, cuenca del río Solo, Java Central
Los fósiles recolectados por los granjeros para von Koenigswald, entre 1937-1941, se conservan en el Instituto Senckenberg de Frankfurt. Se sabe muy poco sobre su procedencia estratigráfica. Entre 1976-1979 trabajó en la zona un equipo japonés-indonesio (Suzuki, 1985). Los fósiles de Homo provienen principalmente de la parte superior de la Formación Sangiran (Pucangan) y la parte inferior y media de la Formación Bapang (Kabuh). Algunas mándíbulas tiene un aspecto masivo, con dientes de aspecto primitivo y de gran tamaño (Kaifu et al, 2005, Kaifu, 2006). Sangiran 2 fue el primer ejemplar recuperado (1937), datados entre 1,51 ma (Larick et al, 2001) y 900 ka (Hyodo et al, 1993).
Los ejemplares más importantes son:
- Sangiran 2, Pithecanthropus II. Descubierto por G.H.R. von Koenigswald en 1937 en Sangiran. La bóveda craneal es muy similar a la del primer cráneo del Hombre de Java, pero menos completa y con una capacidad craneal inferior: sólo 813 cc. 1,5 ma.
- Sangiran 4. 908 cc. 1,6 ma.
- Sangiran 10. 855 cc. 1,2 ma.
- Sangiran 12. 1.059 cc. 1,1 ma.
- Sangiran 17, Pithecanthropus VIII. Descubierto por Sastrohamidjojo Sartono en 1969 en Sangiran. Se trata de un cráneo casi completo, con una capacidad craneal de 1.004 cc. Es el fósil de erectus más completo de Java. Este cráneo es muy robusto. Tienen la cara prominente, muy ancha y bastante plana y los pómulos pronunciados. Sin asociación a instrumentos líticos. 1,3 ma.
- Sangiran 31. Descubierto en 1979. Parte posterior de un cráneo. Paredes muy gruesas y torus occipital muy marcado.
- En 2001, aparecó Bpg 2001.04, un fragmento maxilar (preservando P3-M2), en la Formación Bapang, entre gravas cementadas en la zona Grenzbank. Dos metros por encima del punto del hallazgo, una datación 40Ar/39Ar ha producido una edad de 1,51 ± 0.08 ma. Los patrones dentales sugieren una demo diferente a la representada por los anteriores hallazgos (Zaim et al, 2011).
- En 2016, se recuperó un cráneo adicional, de unos 800 cc.
Ngebung.
- Diente humano junto con una industria lítica muy rica de Modo 1 y numerosos restos de fauna.
- La presencia lítica es una particularidad de este yacimiento ya que en Java los artefactos son raros.
Sambungmacan (0,2 ma)
La morfología de los cráneos es intermedia entre los más antiguos y los más recientes de Ngadong, lo que puede apoyar la hipótesis del aislamiento de las poblaciones de Java.
- Sm-1 (1973). Calota asociada a dos restos líticos. 1.200 cc-
- Sm-2 (1977). Fragmento distal de diáfisis de tibia con morfología moderna.
- Sm-3 (1997). Cráneo. 917 cc.
- Sm-4 (2003). Cráneo. 1.007 cc.
Ngawi.
- Calota de individuo adulto.
Punung
- PU-198, tercer premolar maxilar, de tamaño relativamente pequeño, pero dentro del rango de variación de erectus y sapiens. Datado en hace 143-115 ka. Es preciso tener en cuenta la tendencia hacia la disminución del tamaño de los premolares entre los erectus de Java tempranos y tardíos (Polanski, Marsh y Maddux, 2015).
Los H. erectus asiáticos han sido considerados por algunos autores los únicos H. erectus en sentido estricto. El niño de Mojokerto fue datado por Swisher en más de 1,8 ma y eso significaría que la salida de África es anterior al achelense. Para él, H. erectus evolucionó fuera de África (hipótesis de la salida temprana). Esta hipótesis ha sido puesta en duda por la dificultad de datación. Para Emiliano Aguirre (2008) estos fósiles pertenecen a un grupo descendiente de humanos que salieron de África en el momento de la cladogénesis del H. ergaster. Los ejemplares más arcaicos proceden de Sangiran. Su carácter robusto, especialmente en el aparato masticatorio, justificó la denominación de Meganthtropus palaeojavanicus. Las dataciones de estos ejemplares no son muy diferentes a las de Daminisi, pero sí lo es su morfología (François Sémah, 2014). Al lado de estos restos, aparecen otros más gráciles y no se puede descartar un pronunciado dimorfismo sexual. La mayor parte de los hallazgos son similares al holotipo hallado por Dubois y se clasifican en la denominación Grupo Sangiran-Trinil (Grimaud-Hervé et al, 2012). Fueron recuperados de una capa de sedimentos volcánicos datada en 0,9-0,7 ma (Kabuh layers). Los representantes más derivados proceden de Solo, Sambugmacan y Ngawi (Homo soloensis) y en ellos destaca su encefalización (Grimaud-Hervé, 1997).
- Jacob T (1973) Palaeoanthropological discoveries in Indonesia with special reference to the finds of the last two decades. J Hum Evol2(6), 473-485.
- Holloway RL (1980) Indonesian “Solo”(Ngandong) endocranial reconstructions: Some preliminary observations and comparisons with Neandertal and Homo erectus groups. Am J Phy Anthropol53(2), 285-295.
- Swisher CC, et al (1994) Age of the earliest known hominids in Java, Indonesia. Science263(5150), 1118-1121.
- Swisher CC, et al (1996) Latest Homo erectus of Java: potential contemporaneity with Homo sapiens in southeast Asia. Science274(5294), 1870-1874.
- Anton SC (1997) Endocranial hyperostosis in Sangiran 2, Gibraltar 1, and Shanidar 5. Am J Phys Anthropol102(1), 111-122.
- Antón SC (2001) Cranial evolution in Asian Homo erectus: the Ngandong hominids. Geological Research Development Centre Special Publication 27, 39-46.
- Kaifu Y, Fachroel A, Hisao B (2005) Hominid mandibular remains from Sangiran: 1952–1986 collection. Am J Phys Anthropol128(3), 497-519.
- Dennell RW (2005) The Solo (Ngandong) Homo erectus assemblage: a taphonomic assessment. Archaeology in Oceania40(3), 81-90.
El poblamiento de Europa
El poblamiento de Europa es más reciente que las expansiones de H. erectus hacia Georgia y el Sudeste Asiático. So tres las vías que se han propuesto para la colonización de Europa desde África:
- Gibraltar – El retroceso del nivel del mar durante el avance de los glaciares pudo producir un arco de tierras emergidas entre los Bancos de Fénix en Marruecos y el Bajo de los Cabezos en España. En el Sureste de la Península Ibérica se ha identificado una dispersión de fauna africana (Hippopotamus antiquus, Equus altidens) asociada a la presencia de herramientas olduvaienses, así como Theropithecus oswaldi asociado a tecnología achelense (Ferràndez-Cañadell C, Ribot F y Gibert L, 2015), lo que sugiere que el Estrecho de Gibraltar fue una barrera permeable (Gibert et al, 2015).
- Italia – A través de las islas, aunque no hay evidencias de fauna que pueda haber llegado a Sicilia exclusivamente desde África.
- Corredor de Levante (Bósforo y costa norte del Mar Negro) – Es la vía de salida de África del H. erectus documentada con hallazgos en Israel. En el yacimiento de Ubeidiya (Lago Tiberiades, Israel) aparecieron tres dientes datados en 1,3 ma, asociados a utensilios achelenses de gran formato. Sin embargo, una dispersión transcaucasiana por encima de los 42º de latitud parece difícil debido a las difíciles condiciones climáticas; también el paso a través de Anatolia, cruzando Antitaurus, Taurus, Cárpatos, Alpes y Pirineos.

Bermúdez de Castro y Martinón-Torres (2013) han propuesto un modelo para la evolución del género Homo en Europa durante el Pleistoceno Temprano y Medio basado en la colonización de Europa después de la primera salida de África como resultado de varias migraciones a partir de un área central de dispersiones en Eurasia (CADE), originando demos que migran hacia el este y el oeste del continente euroasiático. Las poblaciones en la CADE y las zonas periféricas se conectarían de forma intermitente debido a la existencia de barreras geográficas y climáticas, con cruzamientos esporádicos entre poblaciones.
- Kozarnika, Bulgaria (1,6-1,4 ma. Sirakov et al, 2010). Molar humano, industria achelense y huesos con incisiones que pueden ser el resultado de un comportamiento simbólico.
- Pirro Nord, Italia (1,6-1,3 ma. Arzarello y Peretto, 2010). Hallazgos líticos.
- Atapuerca y Orce (España).
 La presencia humana parece limitada al sur de la latitud 45ºN, constituida por grupos con tecnologías Modo 1 (Jöris, 2013). Los restos de Atapuerca, datados en 1,3-0,73 ma, demuestran una ocupación temprana de Europa. Quienes eran aquellos Homo y por dónde llegaron, son preguntas difíciles de responder. Quizá derivan de demes europeos de H. erectus, pero serían grupos marginales. Los hombres de la Sima del Elefante se diferencian del H. erectus, pero guardan algunas similitudes con los ejemplares hallados en China. Es posible que procedieran de la misma migración de África hace 1,9 ma. Su viaje por Europa debió recorrer el Corredor de Levante, al mismo tiempo que se adaptaban a las condiciones ambientales locales hasta constituir una población diferenciado que daría lugar al Homo antecessor, pero no hay evidencias de este taxón en Europa aparte de la Península Ibérica.
La presencia humana parece limitada al sur de la latitud 45ºN, constituida por grupos con tecnologías Modo 1 (Jöris, 2013). Los restos de Atapuerca, datados en 1,3-0,73 ma, demuestran una ocupación temprana de Europa. Quienes eran aquellos Homo y por dónde llegaron, son preguntas difíciles de responder. Quizá derivan de demes europeos de H. erectus, pero serían grupos marginales. Los hombres de la Sima del Elefante se diferencian del H. erectus, pero guardan algunas similitudes con los ejemplares hallados en China. Es posible que procedieran de la misma migración de África hace 1,9 ma. Su viaje por Europa debió recorrer el Corredor de Levante, al mismo tiempo que se adaptaban a las condiciones ambientales locales hasta constituir una población diferenciado que daría lugar al Homo antecessor, pero no hay evidencias de este taxón en Europa aparte de la Península Ibérica.Venta Micena, Orce, Granada (1,3 ma)
La expansión del género Homo fuera de África hacia Europa es bastante posterior a la expansión de Homo georgicus y Homo erectus. El material más antiguo encontrado es el denominado Hombre de Orce, con nombre de catálogo fósil VM-0, es un fósil hallado en 1982 por Josep Gibert en el yacimiento de Venta Micena y publicado en 1983, en la provincia de Granada. Varios investigadores propusieron que el fósil era de origen animal en lugar de humano, creando una gran controversia basada en la presencia de una cresta el la cara interna del fósil, lo que sugería que se trataba de restos de un équido y no de un humano. Henry y Marie Antoinette de Lumley fueron los primeros en dudar del Hombre de Orce desde el Musée de l’Homme y el Institut Paleontologie Humaine de París. Sin embargo, el hallazgo, en 2006, de un cráneo de época romana con una cresta semejante a la encontrada en el Hombre de Orce, además de los estudios químicos que demuestran la presencia de albúmina humana, avalan la naturaleza humana del fósil. Sim embargo, estos análisis inmunológicos nunca fueron considerados concluyentes, pues resultaba raro y sospechoso que un fósil tan antiguo conservara cantidades tan grandes de albúmina. Los estudios muestran que la antigüedad del fósil está en torno a 0,9 y 1,6 ma. En la zona se han encontrado piezas manipuladas por el hombre con una datación estimada de 1,4 ma en los yacimientos de Barranco León y Fuente Nueva 3.
Venta Micena
- VM-0, fragmento parietal-occipital, presumiblemente de un niño. Se trata de la parte posterior de un cráneo de un niño de unos 5 años, datado en 1,3-1,2 ma. Para Martínez-Navarro (2002) corresponde a una hembra de bóvido o cérvido. En 2003 se halló en Francolí (Tarragona) el esqueleto de un niño, en una necrópolis del siglo IV, con la misma cresta (Campillo et al, 2006), aunque Martínez-Navarro (2008) afirma que la morfología de las suturas no se parecen. Según el análisis inmunológico efectuado por Concepción Borja et al (1997), el fragmento corresponde a un humano.
- VM-1960, diáfasis humeral juvenil
- VM-3961, parte intermedia de un húmero adulto.
Barranco León D
- BL5-0, parte de un molar, corona y raíz.
- Toro-Moyano et al (2013) sugieren que corresponde a Hippopotamus.
- Francesc Ribot et al (2015) sugiere que perteneció a un humano (parte mesial de un dm2 izquierdo), confirmando la opinión de Arribas y Palmqvist (2002).
- BL02-J54-100, dm1 izquierdo (Isidro Toro-Moyano et al, 2013), de 10 años, Once boy, hallado en julio de 2002, asociado a 1.244 piezas líticas modo 1 y restos óseos con fracturas y marcas de corte. La datación, de 1,4 ma, convierte a este fósil en el resto más antiguo de Europa Occidental. En las excavaciones de 2014 se descubrió un taller lítico.
Francesc Ribot et al (2015) consideran que BL5-0 y BL02-J54-100 podrían corresponder a un mismo individuo.
Otros yacimientos (1,07-0,8 ma)
- Lezignan-la-Cébe (Hérault), Francia hallazgos líticos datados en 1,57 ma.
- Nivel IV del Alto de las Picarazas (Andilla y Chelva, Valencia, España) 1,5 ma, herramientas líticas Modo I en sílex y cuarcita, y además restos de fauna (caballos y bisontes) con marcas de corte. Huesos de conejo con marcas de corte y de dentición humana. De confirmarse la existencia de huesos quemados, sería la evidencia de la producción del fuego más antigua de la Península Ibérica.
- Kurgan Cimbal, Bogatiry y Rodniki, en Rusia. 1,2-0,8 ma (Shchelinsky et al, 2003).
- Valle Medio del Loira, Francia (Despriée et al, 2010). Pont-de-la-Hulauderie, Saint-Hilaire-la-Gravelle. Hallazgos líticos de difícil datación, que podrían tener más de 1 ma.
- Pont-de-Lavaud, Francia, hallazgos líticos datados en 1,1 ma.
- La Chaudronnière
- L’Usine de Rosières à Lunery, Francia, hallazgos líticos datados en 1,1-1 ma.
- Untermassfeld, Alemania, hallazgos líticos datados en 1,07 ma (Landeck y García, 2016).
- Soleihac (Haute-Loire, Francia) indústria lítica 1,07-0,99 ma.
- Grotte du Vallonnet, (Roquebrune-Cap Martin, Alpes Marítimos, Francia) hallazgos líticos datados en 1,05-1 ma (de Lumley et al, 1988).
- Niveles TD3-TD4 de Gran Dolina (Atapuerca) aparecieron algunas herramientas líticas (1 ma). En 2013, se encontró un chopper de calcita tallado.
- Barranco de la Boella (La Canonja, Tarragona), 1 ma, restos líticos y huesos de animales. Entre los restos líticos aparecen dos hachas trabajadas por las dos caras, lo que significaría que estamos ante el achelense más antiguo de Europa.
- El Pino, Carrascosa del Campo, Cuenca, núcleos de cuarcita de cerca de 1 ma.
- Happisburgh, Norfolk, Inglaterra, asentamiento al norte del paralelo 45º. Hallazgos líticos y huellas humanas (Nick Ashton et al, 2014). Datado en 0,99-0,78 ma, cuando la isla estaba unida al continente. Descubierto por Simon Parfitt et al (2010). Para Chris Stringer los autores de la industria quizá pertenecieran al Homo antecessor.
- Dursunlu, Turquía. 0,99-0,78 ma (Gülec et al, 2009).
- Korolevo, Ucrania. 0,99-0,96 Ka (Koulakovska et al, 2010).
- Vallparadís (Terrasa, Barcelona) se han encontrado herramientas modo 1 bipolares olduvaienses datadas en 0,9 ma, con restos de fauna asociados. Algunos restos de herbívoros presentaban marcas de corte y fracturaciones antrópicas.
- Cueva Negra, Caravaca de la Cruz, Murcia, achelense datada en 0,9 ma, con restos de fauna quemada y trozo de sílex deformado por el fuego.
- Cueva Victoria, Cerro de San Ginés de la Jara, a 3 km del Mar Menor y a 13 km al este de Cartagena. Hallazgos datados entre 0,99-0,78 ma. En el mismo lugar se hallaron restos del primate Theropithecus oswaldi. Esta dispersión africana de fauna, junto con la primera presencia del achelense en Cueva Negra apoyan una vía de entrada en Europa por Gibraltar (Gibert et al, 2015). CV-0, falange. CV-1 y CV-2, fragmentos de húmero.
- En Ca’Belvedere di Monte Poggiolo (Forli, Emilia Romagna, Italia), hallazgos líticos datados en 0,85 Ma. Arzarello y Peretto, 2010.
- Dorn-Dürkheim, Alemania, DD31 más de 0,8 Ma. Hallazgos líticos.
- Pakefield, Inglaterra, artefactos de sílex datados en 0,7 Ma.
- En Cúllar-Baza 1 (Granada, España), una escasa colección de piezas de sílex, y en menor medida de cuarcita, que se enmarcan dentro de la tecnología Modo I-Olduvayense datada entre 0,78-0,65 Ma.
- En Solana del Zamborino, Granada, España, industria lítica en su mayor parte realizada con cuarzo y cuarcita aunque también se encuentran instrumentos de sílex, entre los que destaca un gran bifaz desconocido hasta el momento en Andalucía. En 2009 Gary Scott y Luis Gilbert realizaron un estudio geocronológico en el Berkeley Geochronology Center, publicado en la revista Nature, con el que descubrieron que el yacimiento era más antiguo de lo que se creía, situando su cronología de 0,76 Ma.
Sin datación precisa, en la Península Ibérica:
- El Aculadero (Puerto de Santa María, Cádiz).
- El Rompido, Huelva.
- Magoito, Mironço, Aldeia Nova y Açafora, en los alrededores de Lisboa.
- Peteira (Tuy), Tomiño-Campo de Fútbol (Pontevedra), Chan de Vide – “Minas del Condado 3 y 5” (Pontevedra), Montes de Oleiros (Salvaterra de Miño y de As Neves), en las terrazas del tramo final del río Míño.
- Monfarracinos y Toro, en las terrazas del Duero.
- Hallazgos aislados en el entorno de Valladolid, en las terrazas del Pisueerga.
- Perilla del Castro y Betrocino, en las terrazas del Esla.
- Hallazgos en las terrazas del Tormes.
- Pueblanueva, en las terrazas del Tajo.
- Mas d’en Galí (Medinyà, Girona), Costa Roja (Sant Julià de Ramis, Girona) y el conjunto de Puig d’en Roca (Girona), en el curso medio del Ter.
- Gibert, Josep; Agustí, Jordi; Moyà-Solà, Salvador (1983). «Presencia de Homo sp. en el yacimiento del Pleistoceno inferior de Venta Micena (Orce, Granada). Paleontologia i evolució (Publicación Especial 1-9).
- Borja C, García Pacheco JM, García-Olivares E, Scheuenstuhl G, Lowenstein JM (1997) Immunospecificity of albumin detected in 1.6 millionyear-old fossils from Venta Micena in Orce, Granada, Spain. Am J Physi Anthropol 103(4), 433-441.
- Gibert J (2004). El Hombre de Orce los Homínidos que llegaron del Sur. Almuzara Estudios. p. 34.
- Gibert i Clos, Josep, ed. (1999) Los homínidos y su entorno en el Pleistoceno inferior y medio de Eurasia : actas del Congreso Internacional de Paleontología Humana, Orce 1995. Museo de Prehistoria y Paleontología J. Gibert, Ayuntamiento de Orce.
Homo sp.
Sima del Elefante, Atapuerca, Burgos (1,2 ma)
De cronologías similar al Hombre de Orce se encuentra un fragmento de mandíbula encontrado en el yacimiento de Trinchera del Elefante (Atapuerca, Burgos). Datado en 1,2 ma, no se ha atribuido aún a ninguna especie (Homo sp.). Este yacimiento forma parte del complejo de la Trinchera del Ferrocarril en Atapuerca pero no se ha establecido ninguna relación con los materiales encontrados en Gran Dolina o Sima de los Huesos.
- ATE 9-1, en 2007, un fragmento de mandíbula datado en 1,2-1,1 ma. Incluye la región de la sínfisis, donde convergen las dos ramas mandibulares, la derecha conservada hasta el alveolo del primer molar y la izquierda hasta los alveolos del primer y segundo molar. La sínfisis es primitiva. La parte trasera es derivada. El desgaste dental es muy severo. Hipercementosis, infecciones, utilización de palillos para higiene dental, del estudio del cálculo dental se deduce el consumo de plantas y el procesamiento de materias primas vegetales. Los restos de polen y fragmentos de insecto son consistentes con un entorno boscoso. No hay evidencias de cocinado de alimentos (Hardy et al, 2016).
- Un premolar inferior de leche.
- ATE 9-2, Una falange proximal del quinto dedo de la mano izquierda de un adulto. Robusta. (Carlos Lorenzo et al, 2014).
En 2011-2013 se encontraron restos líticos y fósiles animales con marcas de corte de aproximadamente 1,2 ma y en 2013 un fragmento de cuchillo de sílex datado en 1,3 ma. En 2014, se recuperó un lasca de sílex neógeno datada también en 1,3 ma. La tecnología corresponde con el Modo 1.
- Carbonell E, et al (2008) The first hominin of Europe. Nature 452(7186), 465-469.
- Martinón-Torres M, et al (2011) Early Pleistocene human mandible from Sima del Elefante (TE) cave site in Sierra de Atapuerca (Spain): a palaeopathological study. J Hum Evol 61(1), 1-11.
- Bermúdez de Castro JM, et al (2011) Early Pleistocene human mandible from Sima del Elefante (TE) cave site in Sierra de Atapuerca (Spain): a comparative morphological study. J Hum Evol 61(1), 12-25.
- Blasco R, et al (2011) Earliest evidence for human consumption of tortoises in the European Early Pleistocene from Sima del Elefante, Sierra de Atapuerca, Spain. J Hum Evol 61(4), 503-9.
- Lorenzo C, et al (2015) Early Pleistocene human hand phalanx from the Sima del Elefante (TE) cave site in Sierra de Atapuerca (Spain). J Hum Evol 78, 114-21.
- de Lombera-Hermida A, et al (2015) The lithic industry of Sima del Elefante (Atapuerca, Burgos, Spain) in the context of Early and Middle Pleistocene technology in Europe. J Hum Evol 82, 95-106.
- Hardy K, et al (2017) Diet and environment 1.2 million years ago revealed through analysis of dental calculus from Europe’s oldest hominin at Sima del Elefante, Spain. Naturwissenschaften. 104(1-2), 2.
Homo antecessor
Gran Dolina, Atapuerca, Burgos (0,78 ma)
A partir de cata en 1976 se recogen de la Gran Dolina de Atapuerca (Burgos) restos de 0,9 ma (J.M. Parés et al, 2013). El nivel 6, del que proceden los hallazgos, alcanza los 2,5 m de espesor. TD6-2, el estrato Aurora (nombre de la descubridora Aurora Martín Nájera), contenía unos 170 fósiles de Homo antecessor correspondientes a unos once individuos casi todos menores de quince años, la mayoría menores de dos años con algunos bebés. Las dataciones son discrepantes. Falgueres et al (1999) sugieren una datación de hace 850 ka para el estrato Aurora basada en ESR y U-series, estudios biocronológicos y la posición estratigráfica de los hallazgos por debajo del límite geomagnético Matuyama / Bruhnes. Berger et al (2008) utilizando TL e IRSL de alícuotas y análisis paleomagnéticos señaló un intervalo de 950-900 ka.
- ATD6-5 media mandíbula con tres molares de un individuo joven. M3 está saliendo torcida. Sin los rasgos característicos de los preneandertales y pocos rasgos parecidos a los de H. ergaster.
- ATD6-15. Restos parciales de la cara de un niño entre 10 y 11,5 años de edad (Bermúdez de Castro et al, 1997). Conserva el torus derecho y parte de la región de la glabela. Senos frontales muy desarrollados. La capacidad craneal pudo llegar a 1.000 cc.
- ATD6-19. Pequeño fragmento cigomaxilar de un adulto, con un tubérculo cigomaxilar de las mismas características que el de ATD6-58, que se proyecta hacia fuera 2 mm (Arsuaga et al, 1999).
- ATD6-22. Rótula derecha.
- ATD6-38. Muy similar en tamaño y forma a ATD6-69. Su topografía sugiere la presencia de fosa canina (Arsuaga et al, 1999).
- ATD6-43, fragmento de radio casi completo. El índice braquial y las proporciones son más próximos a los del ergaster y a los de los humanos modernos que a los de preneandertales y neandertales.
- El ATD6-50 clavícula muy larga y relativamente delgada, con una curvatura del eje pronunciada y epífisis relativamente pequeñas, similar a la de los neandertales (Carretero et al, 1999).
- ATD6-56. Rótula izquierda asociada a la rótula derecha ATD6-22. Presenta evidencia de un traumatismo.
- ATD6-58. Fragmento cigomaxilar izquierdo de un adulto. Exhibe una notable fosa canina así como un gran tubérculo cigomaxilar situado en posición maxilar que se proyecta hacia fuera 3,3 mm (Arsuaga et al, 1999).
- ATD6-69, El Chico de la Gran Dolina (indviduo 3). Gran parte del lado izquierdo de la cara, excepto las apófisis frontales de los huesos cigomáticos y maxilares. Conserva I2 derecho y P3, M1, M2 y M3 izquierdos permanentes. C y P4 derechos estaban finalizando el proceso de erupción. M2 izquierdo, todavía incluido en su cripta mientras que el germen de M3 se mantuvo unido al hueso por debajo de la cripta de M2. La edad dental, de acuerdo con los estándares modernos, se ha estimado en 10-11 años. Morfología totalmente moderna, incluyendo el patrón de desarrollo dental (Bermúdez de Castro et al, 1999), con rasgos más cercanos a los de los cráneos de Zhoukoudián y Dali y a los de Cromañón que a los neandertales (Rodrigo S. Lacruz et al, 2013, Sarah E. Freidline et al, 2013).
- ATD6-95 y ATD6-70+107 astrágalo izquierdo casi completo y segundo metatarsiano derecho asignados tentativamente al mismo homínido, un macho adulto. El astrágalo es largo y alto, con una tróclea larga y ancha y con un cuello proporcionalmente corto.
- ATD6-96. Fragmento izquierdo de la mandíbula de una mujer muy joven. Exhibe un tubérculo pterigoideo hipertrofiado, una característica incluida en la lista de apomorfías neandertales (Carbonell et al, 2005; Rak et al, 1994; Weaber, 2009).
- ATD6-121. Húmero infantil.
- ATD6-124. 3/4 partes de un cuarto metatarsiano derecho. Según L. Martín-Francés et al (2013) el hueso estaba roto y se estaba soldando en el momento del fallecimiento del individuo. La presencia de esta fractura sugiere que la correspondiente población recorría grandes distancias o se desplazaba frecuentemente por un terreno desigual.
- ATD6-148. Rafa. Húmero izquierdo de adulto muy joven vigoroso. Quebrado para obtener la médula. Marcas de descarnado y destrozo de los cóndilos para separar las masas musculares. Al igual que ATD6-121, exhibe una gran fosa del olécranon y unos pilares laterales y mediales muy finos, similares a los de los homínidos del Pleistoceno Medio europeo, los neandertales y Bodo (Carretero et al, 2009; Bermúdez de Castro et al, 2012).
- ATD6-100/168. Un gran fragmento de parietal, el mayor recuperado hasta el momento del neurocráneo de antecessor. La morfología de las fracturas es compatible con un trauma peri-mortem. El hueso perteneció a un individuo juvenil (Bruner et al, 2016).
Además, más de 800 artefactos líticos: En el nivel TD4, un chopper que fue utilizado para descuartizar huesos de animales, datado en 1 Ma. El techo del nivel TD4 había deparado ya varias herramientas de cuarcita manufacturadas a partir de cantos del río Arlanzón. En TD5, tres artefactos, entre ellos un núcleo de cuarcita y una lasca de sílex. En TD6, asociada a los restos humanos, aparece una abundante tecnología lítica con núcleos y lascas (industrias arcaicas o Modo I); la materia prima utilizada es el sílex, cuarzo, cuarcita, caliza y arenisca.
- Plesiomorfías:
- La dentición: desarrollo de un cíngulo en premolares y molares, crenulaciones del esmalte profusas, asimetría de la corona del primer premolar inferior, gran tamaño en molares y sistemas radiculares complejos y robustos.
- Los rasgos endocraneales de los parietales.
- Frente huidiza y toro supraorbital de doble arco.
- Ausencia de mentón.
- Apomorfías:
- Capacidad craneal grande, estimada en más de 1.000 cc.
- La cara más moderna que la de los honininos africanos del Pleistoceno Inferior, caracterizados por un prognatismo pronunciado. Es plana, con un nasal saliente, maxilar poco proyectado y mandíbula delgada.
- El patrón de remodelación facial en la ontogenia.
- La ontogenia dental.
- Comparte con los neandertales y los HAM un borde superior convexo de la escama del temporal, y una posición anterior del conducto incisivo, que es casi vertical (Arsuaga et al, 1999).
- La región mastoidea es una reminiscencia de la de los neandertales.
- Fosa canina, consecuencia de una reorientación hacia delante y abajo de las placas infraorbitarias.
La atribución taxonómica de estos fósiles es dificil. Lo habitual para los restos europeos de transición era clasificarlos como H. heidelbergensis. Pero Carbonell (1995) sugiere que se trataría de un taxón diferente a H. erectus (que se extingue) y al H. ergaster (que daría lugar a H. sapiens). Howell le asignó el paleodemo Atapuerca-Gran Dolina. El equipo de Atapuerca propuso la nueva especie Homo antecessor (Carbonell et al, 1997) con una topografía mediofacial completamente moderna y otros rasgos que lo apartan de H. erectus, H. ergaster y H. neanderthalensis.
Homo heidelbergensis
Hallazgos en España
Sima de los Huesos, Atapuerca (0,427 ma)
En la Sima de los Huesos (Atapuerca, Ibeas de Juarros, Burgos, España) se hallaron fósiles del Pleistoceno Medio de unos 28 individuos: un niño, doce adolescentes, diez subadultos, y diez adultos de más de 25 años de los que solo tres superarían los 35 años. Ninguno superaría los 45. Ambos sexos están representados, con una diferencia de tamaño de un 10%. La datación del yacimiento has sido objeto de controversia. Las diferentes estimaciones fluctuan entre 0,4 y 0,5 ma. Los primeros restos fueron descubiertos en 1976 por el equipo de Trino Torres. Lee J. Arnold et al (2014) obtuvieron una datación mínima de 427 ± 12 ka. El análisis del ADN nuclear ha confirmado la pertenencia de esta población al linaje de los Neandertales (Meyer et al, 2016).
Para Emiliano Aguirre (2008) estos individuos se hallaban juntos en el interior de una cueva cuya entrada se cerró accidentalmente. Los restos se deslizaron hasta la boca de la sima y los carnívoros dejaron su huellas en los huesos. Otra interpretación más moderna identifica la Sima de los Huesos como un cementerio. Arsuaga, cree que aquellos homínidos fueron arrojados deliberadamente. Nohemi Sala et al (2014) han descartado la acumulación por carnívoros. Según Aranburu, Arsuaga y Sala (2015) la acumulación de homínidos se produjo en un único evento, sin transporte. En el momento de la acumulación de fósiles el único acceso posible a la cámara era un conducto vertical profundo, imposible de escalar (la chimenea C2 que conecta al oeste con la Sala de los Cíclopes SCi y al este con un conducto bajo e inclinado OC).
Se han descubierto 17 cráneos (Arsuaga et al, 2014) muy similares entre sí y distinguibles de los de otras poblaciones europeas de datación similar. El patrón morfológico es consistente con características derivadas de los Neandertales, tanto en la cara como en la bóveda craneal, muchas de las cuales están relacionadas con el aparato masticatorio incluyendo su uso como tercera mano, con gran desgaste de los incisivos. Esta población se situaría en las base de la evolución de los Neandertales. La modificación facial fue el primer paso, dentro de un patrón en mosaico. Entre las características que les aproxima a los Neandertales están la forma triangular de la cara y los arcos superciliares, prognatismo, mandíbulas inferiores robustas, patrones de las cúspides dentales y molares pequeños. Algunos investigadores sugieren que taxonómicamente no es posible clasificar esta población dentro de H. heidelbergenesis, pero tampoco en H. neanderthalensis, y se incluirían dentro de los preneandertales, junto con Reilingen, Steinheim y Swanscombe. Tattersall (2011) considera que los restos de la Sima de los Huesos son de filiación desconocida, al igual que los de Steinheim y Reilingen. Para Martinón-Torres y Bermúdez de Castro (2011) la morfología de los dientes de Sima de los Huesos no difieren de los de los Neandertales.
- 17 cráneos (Arsuaga et al, 2014) similares entre sí y distinguibles de los de otras poblaciones europeas de datación similar, lo que indica que estas poblaciones pertenecían a diferentes demos.
- 20 mandíbulas (medio centenar de piezas), cuatro de ellas enteras. Son robustas, sin mentón. Cuerpo mandibular bajo, con el borde inferior ancho, plano, paralelo a la fila de dientes y con un acusado reborde. Ramas ascendentes largas y con el borde superior poco escotado. En seis individuos se observa una artrosis degenerativa de la articulación del hueso temporal y la mandíbula.
- Más de 500 dientes.
- 42 restos de clavículas, 114 de escápulas, 645 restos de vértebras, 466 de costillas con un cuerpo ancho, una plesiomorfia heredada (Arsuaga et al, 2015).
- 110 restos de húmeros, 65 de cúbitos y 89 de radios (49 de adultos).
- 173 restos de fémures, 20 de rótulas, 89 de tibias y 105 de peronés. Los huesos largos son muy fuertes.
- 121 huesos del carpo, 103 del tarso, 93 metacarpianos, 80 metatarsianos, 208 falanges de pies y 304 falanges de manos. En los restos de niños, se dan varios casos de líneas de Harris, consecuencia típica de enfermedades infantiles.
- Hioides y huesos del oído. Ignacio Martínez ha deducido de su análisis la capacidad de habla de estos individuos.
- Pelvis 1 (Elvis). 0,5 ma. Pelvis con fuertes inserciones musculares. Es la pelvis más ancha y mejor conservada de todo el registro fósil, junto con otros restos del esqueleto, correspondiente a un varón muy corpulento que padeció importantes enfermedades degenerativas (espondilolistesis) desde mucho antes de morir, con más de 45 años de edad. Debió medir entre 1,70-1,75 m y pesar 90-95 kg.
- Agamenón, Atapuerca 4. Caja craneal completa, sin la cara. Se observa una artritis avanzada en los cónditos occipitales y los dos conductos auditivos obstruidos por hiperostosis. El frontal está inclinado, la bóveda craneal baja y el occipital alargado.
- Miguelón, Atapuerca 5 (AT 700). Descubierto entre 1992 y 1993. Su antigüedad se estima en 0,3 ma; su capacidad craneal es de 1.125 cc (Arsuaga et al. 1993; Johanson and Edgar 1996). Se ha identificado la mandíbula que le corresponde.
- Benjamina. Individuo de 10 años de edad. Padeció craneosinostosis, enfermedad que produce una sutura anticipada de los huesos craneales, lo que se traduce en un retraso psicomotor.
- AT 624. Individuo infantil con un fuerte golpe en el lateral del torus supraorbitario izquierdo, con signos de regeneración ósea.
- En 2014 se recuperaron restos del coxal derecho de un adolescente de unos 13 años, una vértebra de otro individuo adulto y un occipital, y un diente de otro homínido de unos 50 años, así como varios huesos de la nariz, manos y pies.
- En 2015, restos craneales adicionales.
Galería, Atapuerca (0,3 ma).
- AT 76-T1H. Fragmento de mandíbula derecha con dos molares, hallado por Trino Torres (1976). La presencia de espacio retromolar, el toro mandibular y el tamaño de los últimos molares jutifican su asignación al demo de los preneandertales europeos.
- Fragmento de cráneo hallado en 1995 que comprende la parte posteromesial del parietal derecho, parte posterior de la sutura sagital, y otro fragmento menor del parietal izquierdo, con una porción muy pequeña adherida del occipital que incluye el lambda. El parietal es muy grueso y fue hundido por un golpe que luego sanó.
- Restos líticos Modo 2.
- Del análisis polínico se desprende un periodo relativamente frío.
Gran Dolina, Atapuerca (0,45 ma)
- En el nivel TD10 apareció indústria lítica del Modo 2 asociada a un procesamiento sistemático y direccional de carcasas, incluyendo eventos de caza, datado en 0,45 ma. TD10-1 puede representar la evolución local desde el Modo 2 al Modo 3 (Andréu Ollé et al, 2013).
Pinilla del Valle, Madrid: 2 dientes
- De momento sólo se han encontrado 2 dientes, probablemente de Neandertal..
Sima de la Cueva del Ángel, Lucena, Córdoba
- Un fémur. Los trabajos que se desarrollaron en la Cueva del Ángel, en los años 1995, 1996, 2002 y 2003, permitieron detectar la existencia de un potente depósito arqueológico con una gran amplitud cronológica que se situaría entre los 0,48-0,1 ka. Podría constituir uno de los últimos lugares de ocupación de H. heidelbergenesis, en un momento en que los Neandertales poblaban Europa.
Cueva de Lezetxiki, Mondragón, Guipúzcoa
- Húmero completo de un individuo femenino. Datación incierta. En 2014 fue datado directamente en un mínimo de 165 ka.
Tossal de la Font de Vilafamés, Castellón
- Fragmento distal de húmero y fragmento de coxal.
Bolomor, Valencia
- Varios dientes y fragmento de peroné.
Valdegoba, Burgos
- Mandíbula, dientes y tres restos poscraneales.
Cueva de Mollet, Parc de les Coves Prehistòriquess de Serinyà, Pla de l’Estany, Girona
- Un diente de infante, datado en 215 ka.
Conca de Dalta, Lleida
- En febrero de 2013, se publicó la noticia del hallazgo en una cueva de restos neandertales y herramientas achelenses, datados en 200 ka.
Hallazgos en Italia
- En Saccopastore, cerca de Roma, aparecieron entre 1929-1935 un cráneo humano masculino y otro femenino, de características arcaicas, similares a los cráneos de la Sima de los Huesos (Arsuaga el al, 1997). Según una reciente revisión estratigráfica de Marra et al (2015) los cráneos se depositaron hace ca 250 ka, en el inicio de MIS 7. Saccopastore 1 está casi completo y muestra arcos superciliares muy grandes, frente inclinada baja y bóveda occipital redondeada. Los inicios del tercio medio facial son amplidos, con grandes dientes y nariz.
- En Notarchirico (Basilicata) aparecieron restos líticos Modo 2 datados en 0,6 ma. En la base hay restos de industrias de Modo 1.
- En La Pineta di Isernia fue hallado en 2014 un incisivo deciduo (IS42) en el nivel arqueológico 3 coll (que recubre la capa 3a). Se trata del resto humano más antiguo de Italia, datado por 40Ar/39Ar, entre hace 583-561 ka, a finales de MIS 15 (Peretto et al, 2015). Está asociado a herramientas no achelenses, pese a ser un poco más joven que la difusión del achelense en Europa Occidental, lo que apoyaría la continuidad en refugios de las poblaciones europeas preexistentes.
- En 1994 Italo Bidditu halló entre los sedimentos de una cuneta de una carretera en Ceprano, Frosinone, un cráneo que conserva prácticamente toda la bóveda, pero sin base, cuya capacidad podría ser hasta de 1.200 cc. En un principio fue datado en más de 800 ka, pero una revisión de Aurélien Mounier et al, 2011 lo data en 0,430-0,385 ma. No está asociado a macro o microfauna, restos polínicos ni industria lítica. Debido a la datación dudosa y a sus características peculiares, fue relacionado con muy diferentes posiciones filogenéticas, e incluso le fue asignada una nueva especie: Homo cepranensis (Mallegni et al, 2003), pero actualmente se asigna a H. heidelbergensis (Manzi, 2016).
- Fontana Ranuccio. Dientes datados en 0,458 ma: dos primeros molares inferiores (izquierdo y derecho), un incisivo inferior izquierdo y un posible canino superior izquierdo. La comparación con otros ejemplares contemporáneos muestra que los dientes de Fontana Ranuccio son morfológicamente similares a Atapuerca-Sima de los Huesos, Arago XIII y neandertales (Mauro Rubini et al, 2014).
- Cava Pompi. Fragmentos de cráneo, cúbito y tibia. 0,45 ma.
- Visogliano. Mandíbula y cinco dientes sueltos. 0,4 ma.
- Vanosa-Notarchirico. Fémur asociado a industria de Modo 2. No se ve técnica de Levallois. 0,4-0,35 ma.
- Castel di Guido. Fragmentos craneales. Una diáfasis de fémur. Asociados a industria Modo 2. 0,4 ma. (Alciati et al, 2005; Capasso, Michetti y D’Anastasio, 2008). El espécimen CdG-1 fue considerado inicialmente un fragmento de hioides, pero según el estudio de Capasso et al (2016), se trata de un borde posterior de la primera vértebra cervical.
- Ponte Mammolo. Un fémur. 0,2 ma.
- Grotta del Principe. Pelvis de individuo femenino. 0,2 mda.
- Sedia del Diavolo. Fragmentos de cráneo y fémur y un metatarsiano. 0,2 ma.
- Grotta del Poggio. Un diente y varios poscraneales. 0,2 ma.
- Casal de Pazzi. Fragmento de parietal. 0,2 ma.
Estos hallazgos sugieren la presencia de refugios para la población humana en el centro sur de Italia hasta, al menos, el periodo MIS 11 (Peretto et al, 2015).
Hallazgos en Alemania
- Steinheim (Stuttgart). 1934. Cráneo casi completo, deformado y restaurado. Unos 1.100 cc. Largo y estrecho. Moño occipital y apomorfias de neandertal poco avanzadas, pero la cara no muestra rasgos neandertales. 0,225 ma (Adam, 1954, 1985). Por su pequeño tamaño, posiblemente corresponda a una hembra. Caja craneana bastante esférica. Órbitas relativamente pequeñas, coronadas por prominentes rebordes supraorbitarios en doble arco. Orificio nasal bajo y ancho. Pómulos retraídos formando una pequeña fosa y no engrosados como en los neandertales. Fosa canina.
- Ehringsdorf. Restos craneales y poscraneales de varios individuos. Rasgos de neandertal arcaico. 0,25-0,2 ma.
- Reilingen. Cráneo incompleto. Algunos rasgos afines a los Neandertales.
- Heidelberg, Mauer Jaw, Descubierto por trabajadores de una cantera en 1907, cerca de Heildelberg. Descontextualizado de su posición estratigráfica, su edad es incierta, pero se ha estimado en hace 609 ka (Wagner et al, 2010). Se trata de una mandíbula inferior, ramas ascendentes muy anchas, con el mentón retraído y todos los dientes. Escotadura sigmoideia apenas deprimida, sínfisis mandibular alta y retraída. Molares relativamente pequeñas, sobre todo M3. La mandíbula es extremadamente larga y robusta, como la del Homo erectus, pero con los dientes más pequeños. Es muy difícil interpretar el espécimen desde una perspectiva filogenética.
- Bilzingsleben (Valle Wipper, Turingia). Fragmentos de al menos tres individuos, asociados a industria de Modo 1 evolucionado (micro-Clactoniense) y fauna. Se han reconstruido dos cráneos, sin rasgos neandertales. Vlcek (1989) y Mania et al (1994) los asignan a H. erectus. 0,3-0,28 ma (Mania y Thomae, 2006). Ambiente de bosque con un clima ligeramente más templado que el actual. Aparecieron más de 10.000 huesos de animales, algunos claramente grabados.También tres estructuras de hábitat y organización del espacio.
- Schöningen (Elba-Saale) contiene al menos 20 sitios arqueológicos de preservación excepcional en un área de unos 9.400 m2 (Serangeli et al, 2015), con una sucesión sedimentaria de hasta 45 m de espesor (Lang et al, 2015). Schöningen 13/II sedimentary sequence 4 (Spear Horizon) es conocido por sus importantes hallazgos. En 1995 se descubrieron numerosos artefactos de madera y los esqueletos de unos 20-25 caballos descuartizados. No está clara la forma en que los materiales fueron depositados (Mareike C. Stahlschmidt et al, 2015). Se ha estimado una datación máxima de 337-300 ka (Daniel Richter y Matthias Krbetschek, 2015). Indicios de fuego (Mareike C. Stahlschmidt et al, 2015; Utz Böhner, Jordi Serangeli y Pascale Richter, 2015). Nueve lanzas (de empuje), una jabalina (para lanzar), un palo con punta por ambos lados y un palo puntiagudo quemado, obtenidas del tronco de una pícea, dándoles forma con cuidado, tallando las puntas en la madera de la base del tronco, particularmente dura (Schoch et al, 2015).
Hallazgos en Reino Unido
- Swanscombe (East Anglia, Reino Unido). Occipital muy completo (1935), parietal izquierdo (1936) y derecho (1955) de hembra joven adulta. 1.300 cc. Paredes gruesas. Occipital redondeado. Ambas características lo asemejan a los neandertales, pero la opinión no es unánime. 0,3-0,4 ma (Stringer y Hublin, 1999). Gran número de útiles líticos achelenses, incluyendo bifaces, raspadores y denticulados.
- Hoxne, Clacton-on-Sea, Barnham e High Lodge (East Anglia, Reino Unido). Registro fósil y de industria Modo 2. Cultura clactoniense.
- Pontnewydd (Norte de Gales). Compton y Stringer (2015). Fragmentos de maxilar y tibia. 17 dientes. Hallados entre 1980-1995. 0,251-0,195 ma. Asociados con hachas de mano y artefactos levallois.
- Boxgrove (East Anglia). Excavado por Mark Roberts en los 90. Se han encontrado huesos de elefante, rinoceronte, caballo, ciervo, león, lobo, hiena, castor, liebre, ratón de campo y rana. Muchos huesos conservan marcas líticas. Los únicos restos humanos son dos dientes y parte de una tibia, grande y robusta, que corresponde a un individuo de 1,80 m y 90 kg (0,5 ma); su morfología es similar a la de los neandertales. Los espeleotemas fueron datados por U-series de alta resolución en 600 ka (Bischoff et al, 2007), pero la validez de esta datación para los restos humanos es discutible (Endicott et al, 2010).
Hallazgos en Francia
- Montmaurin (Francia). Mandíbula. Rasgos de preneandertal.
- Biache-Saint Vaast. Tres fragmentos de cráneo atribuidos al mismo individuo. Se observan rasgos de los neandertales clásicos. Trazado de la red meníngea avanzado, parecido a Swanscombe y con Neandertales.
- Biache 1. Asociado a abundante industria musteriense tipo La Ferrasie. Solo se preserva la mitad posterior, que presenta morfología neandertal.
- Biache 2, fragmentado y mezclado con fauna. Muy robusto.
- Tourville-la-Rivière. Tres huesos largos de la misma extremidad superior izquierda de un joven o adulto, descubiertos en 2010 por A. Cottard y A. Thomannde en la capa D2inf del yacimiento al aire libre de Tourville-la-Rivière en el valle del Sena en el norte de Francia, donde reposaron tras un transporte fluvial postdeposicional. Las dataciones proporcionan un rango de edad de 226-183 ka (finales de MIS 7). Jean-Philippe Faivre et al (2014) atribuyen los restos humanos a Homo neanderthalensis, aunque de forma no segura.
- Tauvatel, Arago (Taltaüll, Pirineos Orientales). Yacimiento con veinte niveles de ocupación entre 0,55-0,34 ma, excavado bajo la dirección de Henry y Marie-Antoinette de Lumely. 80 fósiles en el suelo G, producto del canibalismo, datados en 0,45 ma. Se mezclan características primitivas con otras típicas de los neandertales, incluso en grado más derivado, si bien en Arago 2 y Arago 13 están ausentes las características derivadas neandertales; si bien su robustez es diferente, ambas guardan similitudes con la mandíbula de Mauer.
- Arago 21, Tautavel Man. Descubierto en 1971 por Henry de Lumley en el estrato III. Rostro casi completo, con 5 dientes molares y parte de la bóveda craneal. Su capacidad craneal es de 1.150 cc. Bóveda craneana baja, huesos malares y rostro macizos, pómulos engrosados, maxilar inflado, sin fosa poscanina; parte media de la cara proyectada hacia delante; el frontal es estrecho, la angulación del occipital acusada, parietal grueso y corto; las líneas temporales son altas y marcadas; órbitas relativamente bajas y cuadrangulares, coronadas por fuertes rebordes supraorbitales en doble arco, con depresión glabelar.
- Arago 2. Mandíbula de un individuo femenino. Prognatismo.
- Arago 13. Mandíbula de un individuo masculino.
- Dientes de 10 individuos distintos.
- Arago 44. Hueso ilíaco. En conjunto con otros huesos nos permite estimar una talla de 1,64 m.
- Arago 131. Mandíbula con dos dientes, descubierta en 2012.
- Macromamíferos y herramientas líticas.
- En 2013 se hallaron siete dientes, una mandíbula y un fémur.
- En 2015, Arago 149, un incisivo central inferior muy desgastado, en un nivel de ocupación datado en 0,56 ma.
- Vergranne, canino de menos de 0,5 ma.
- Pech-de-l’Aze. Infantil. 0,4-0,2 ma.
- Orgnac 3. Una decena de dientes aislados, parecidos a los de Arago. Aparece técnica Levallois. 0,35-0,32 ma.
Hallazgos en Grecia
- Petralona 1. Cráneo casi completo hallado casualmente en 1959, literalmente pegado a la pared de la cueva y cubierto por una capa de calcita. El resto del esqueleto había desaparecido. Presenta apomorfias neandertales en órbitas, rostro, frente y bóveda craneanas, con una cara alargada y plana y una mandíbula muy ancha. Su capacidad craneana es de 1.220 cc. Su edad se estima en 0,25-0,15 ma (Grun, 1996). Grandes rebordes supraorbitales en forma de doble arco y grandes órbitas de forma aproximadamente cuadrangular. Pómulos hinchados y maxilar inflado como en los Neandertales.
- Apidima I y II. Dos cráneos. 0,2 ma. Rostro, frente y bóveda craneal muy próximos a los Neandertales de Saccopastore y Gibraltar.
- Marathousa. Artefactos líticos asociados con restos de elefante con marcas de corte. Datado entre 0,6-0,3 ma.
Hallazgos en Hungría
Vérteszöllös. 1965. Occipital y molar deciduo, asociados con fauna e industria clactoniense. 0,475-0,25 ma (Cherdyntsev, 1971). El occipital no se parece al de los neandertales y es más primitivo (Thoma, 1966).
- Kokkoros, P., and A. Kanellis. «Découverte d’un crane d’homme paléolithique dans péninsule Chalcidique.» Anthropologie 64 (1960): 132-147.
- Stringer, Christopher B., F. Clark Howell, and John K. Melentis. «The significance of the fossil hominid skull from Petralona, Greece.» Journal.of.Archaeological.Science 6.3 (1979): 235-253.
- Hennig, G. J., et al. «ESR7dating of the fossil hominid cranium from Petralona Cave, Greece.» (1981): 533-536.
- Poulianos, Aris N. «Petralona cave dating controversy.» (1982): 280-280.
- Rightmire, G. Philip. «Patterns of hominid evolution and dispersal in the Middle Pleistocene.» Quaternary.International 75.1 (2001): 77-84.
- Beck, Curt W. «Archaeometric Clearinghouse.»Journal of Field Archaeology 9.2 (1982): 279-280.
- de Lumley H, de Lumley MA (1971). Découverte de restes humains anténéanderthaliens datés du début du Riss à la Caune de l’Arago (Tautavel, Pyrénées Orientales). Comptes Rendues des seances de l’Acacdemie de Science de Paris 272, 1729-1742.
- de Lumley H, de Lumley MA (1973) PreNeanderthal remains from Arago cave in
southwestern France. Yearb. Phys. Anthropol 17, 162-168.- Yokoyama Y, Nguyen HV (1981) Direct dating by non destructive gamma ray spectrometry of fossil human skull Arago XXI, fossil animal bones and stalagmites of the Caune de l’Arago at Tautavel. Proceedings Pretirage, 355-375.
- Rightmire GPh (2001) Patterns of hominid evolution and dispersal in the Middle Pleistocene. Quaternary International 75(1), 77-84.
- Beck CW (1982) Archaeometric Clearinghouse. J Field Archaeol 9(2), 279-280.
- Falguères Ch, et al (2004) New U series dates at the Caune de l’Arago, France. J Archaeol Sci 31(7), 941-952.
- Byrne L (2004) Lithic tools from Arago cave, Tautavel (Pyrénées Orientales, France): behavioural continuity or raw material determinism during the Middle Pleistocene? J Archaeol Sci 31(4), 351-364.
Homo rhodesiensis / heidelbergensis
- Nadung’a, lado oeste del Lago Turkana. Molar datado en 750 ka, el de tamaño más pequeño conocido en África del Pleistoceno Medio (Maddux et al, 2014).
- Ternifine (Argelia) >0,6 Ma.
- Bodo (Etiopía). Quizá el más antiguo, descubierto entre 1976-1981. 0,6-0,4 ma (Clark et al, 1994). Cara casi completa y parte de las paredes laterales. Muy robusta. Orificio nasal más ancho de todo el registro fósil humano. Asociado a herramientas del achelense final.
- Thomas Qarry I. Parte de una mandíbula (Ennouchi, 1969). Entre 1994 y 2011, se recuperaron nuevos fósiles de la unidad estratigráfica 4. Una diáfisis de fémur, varios dientes aislados, fragmentos craneales, vértebras, una mandíbula completa de un adulto y una mandíbula parcial de un niño de corta edad. ca 0,5 ka. Los artefactos achelenses recuperados responden a una producción lítica en la propia cueva. Rica en restos de fauna de mamíferos acumulados por carnívoros, incluyendo Homo. Daujeard et al, 2016.
- Thomas Qarry III. Fragmento craneal y varios dientes, asociados a industria achelense. 0,4 Ma.
- Salé (Marruecos). Cráneo. 0,4 Ma.
- Ndutu (Tanzania) descubierto en 1973. Fragmentos craneales relativamente delgados por lo que pudieron pertenecer a una hembra.
- Olduvai OH11, OH22, OH23 (Tanzania). 0,3 Ma.
- Eyasi 1 y Eyasi 2 (Tanzania) descubiertos entre 1935-38. Domínguez-Rodrigo et al, 2007, 2008; Mehlman 1987).
- Laynyamok. Tres dientes y un fragmento de fémur.
- Ileret, Kenia. KNM-ER 3884 cráneo descubierto el 1976. Fémur ER 999.
- Hopefield 1 (Saldanha, Elandsfontein, en Africa del Sur) descubierto el 1953. 27 fragmentos de la bóveda craneal, asociados a una mandíbula, industria lítica y fauna. Bóveda muy robusta, gruesa, aplastada, con relieve supraorbital continuo y poderoso y repliegue occipital prolongado. El perfil de la región occipital es anguloso. 0,3 Ma.
- Hoedjiespunt, África del Sur. 0,3-0,2 Ma.
- Cueva Gladysvale en el Valle Sterkfontein, Sudáfrica. Phillip Taru y Lucinda Backwell (2013) hallaron pelos fósiles humanos en coprolitos de hienas marrones (Parahyaena brunnea) datados en 257-195 ka.
- Kabwe 1 (Zambia). El hombre de Broken Hill. 0,4 Ma, pero la datación es difícil. Holotipo de Homo rhodesiensis Woodward 1921. Cráneo completo de gran robustez, torus supraorbital fuerte, pliegue occipital saliente. Frente deprimida, rosto prognato. 1.280 cc de capacidad craneal. Las paredes del cráneo son subverticales y no covergen con claridad hacia arriba. Paredes gruesas, pero no tanto como en erectus. Protuberancias parietales salientes. Occipital sensiblemente inclinado hacia atrás en un ángulo menos cerrado que el de erectus. Grandes órbitas oculares de contorno algo cuadrangular con grandes arcos superciliares pero no continuos como en erectus. Enorme orificio nasal triangular. En el mismo yacimiento se hallaron otros dos fragmentos craneales, trozos de un maxilar y varios del esqueleto poscraneal incluyendo una tibia completa, fémur, pelvis y sacro, datados entre más de 0,3-0,21 Ma.
Las características craneales recuerdan las de los neandertales, pero las proporciones corporales son más parecidas a las del sapiens.
- Clark JD, et al (1947) New studies on Rhodesian man. The Journal of the Royal Anthropological Institute of Great Britain and Ireland 77(1), 7-32
- Klein RG (1973) Geological antiquity of Rhodesian man. 311-312.
- Stringer CB (1986) An archaic character in the Broken Hill innominate E. 719.» Am J Phys Anthropol 71(1), 115-120
- Rightmire PG (1996) The human cranium from Bodo, Ethiopia: evidence for speciation in the Middle Pleistocene? J Hum Evol 31(1), 21-39.
- Carretero JM, et al (2008) A partial distal humerus from the Middle Pleistocene deposits at Bodo, Middle Awash, Ethiopia. Anthropological Science : 0807280042.
Homo neanderthalensis
Hay muchos yacimientos con presencia de Neandertales en Europa y Asia, poro no en África. Los fósiles de la última parte del Pleistoceno Medio (desde hace unos 200.000 años) ya pueden ser considerados verdaderos y completos neandertales. A partir de hace 127 ka, los fósiles de neandertales empiezan a ser abundantes en los yacimientos europeos, y se encuentran también en Asia central y Oriente Próximo, lugares a donde emigraron desde Europa. Fósiles tan emblemáticos como Le Moustier, que da nombre al Musteriense, Guattari 1 (Monte Circeo) o La Chapelle-aux-Saints, vivieron en Europa hace menos de 60.000 años. Estos neandertales tienen algunas características nuevas con respecto a los anteriores, y son llamados a menudo «neandertales clásicos». Los últimos neandertales vivieron en Europa hasta hace unos 30.000 años, e incluso algo menos, antes de extinguirse.
Anteriores a 134 ka
Italia
- Cueva de Lamalunga (Martina Lari et al, 2015), esqueleto hallado en 1993 en el karst de la Cueva de Lamalunga, cerca de Altamura, en el sur de Italiam datado entre 172 ± 15 ka a 130,1 ± 1,9 ka.
España
- Cueva Bajondillo (Málaga), evidencias de neandertales y del consumo de marisco, con una datación de 150 ka (Miguel Cortés-Sánchez y otros, 2011).
- Tossal de la Font (Castellón), un fragmento de húmero y otro de pelvis.
Francia
- Le Lazaret. Paietal de inmaduro, con cicatriz de trauma y meningioma postraumático; un incisivo lacteal y un canino adulto. Asociado a industria Modo 2 evolucionado. 0,16 ma.
- Grotte Suard. La Chaise. Partes de un cráneo adulto, otro infantil y su mandíbula, tres dientes. Asociados a industria Modo 2 evolucionado, con lascas Levallois. 0,13 ma.
- Orgnac 3 (Combier, 1967). Siete dientes asociados a unos pocos bifaces.
- Payre G y F (Moncel et al, 2002). Parietal y ocho dientes asociados a unos pocos bifaces.
- Abri Moula c. XVI. Un infante.
Datados en OIS5 (134.000-74.000)
Próximo Oriente
- Tabün. Pelvis de mujer neandertal muy completa.
Francia
- Cueva de Crombe Grenal, Dordoña, Francia. Ocupada por neandertales entre 115-50 ka. Excavada en 1950-1960 por François Bordes. Según el estudio de Phillip Chase de los huesos de animales hallados en el yacimiento, los neandertales cazaban ciervos y renos, de uno en uno, y aprovechaban la carroña de caballos y bóvidos. Indicios de utilización simbólica del esqueleto de rapaces diurnas (Morin y Laroulandie, 2012).
- Abrigo Bourgeois-Delaunay. Hueso temporal primitivo.
- Moula-Guercy. Fragmentos craneales. Restos poscraneales.
- Artenac (Charente). Fragmentos craneales.
- Abri Moula c. XV (Defleur et al 1999). 90 restos humanos.
- Payre E y D. Dos dientes.
- Abri des Pêcheurs. Siete dientes.
- Abri du Maras, nivel 5.
- La Chaise. El sitio se compone de tres cuevas: Bourgeois-Delauny, Suard y Duport. Descubierta en la década de 1850. De particular importancia es el Abri Bourgeois-Delauny que contenía los restos de 23 fósiles de adultos e infantil. El material fósil se compone principalmente de un cráneo parcial, fragmentos de mandíbula y dientes. El material del Abri Suard consiste en 52 adultos fragmentarios y bebés. Sobresalen una gran parte de una bóveda craneal, un hueso occipital, fragmentos de mandíbula y dientes asociados.
Alemania
- Taubach.
- Salzgitter.
- Ehringsdorf. Cráneo.
Italia
- Grotta Guattari (Monte Circeo). Un cráneo y dos mandíbulas. Afectado por la acción de las hienas.
Eslovaquia
- Gánovce. Fragmentos de cráneo, molde endocraneano y otros de poscraneanos.
Grecia
- Acantilado de Apidima. Apidima I, medio cráneo. Apidima II, varios fósiles humanos con partes del cráneo facial. Primitivos.
Croacia
- Kaprina, datado en 130 ka, contiene cientos de fósiles de unos ochenta individuos. Es un refugio de roca arenisca descubierto por Dragutin Gorjanovic-Kramberger en 1899 cerca de Krapina (Croacia). Excavado entre 1899-1905, este yacimiento proporcionó también más de 800 herramientas líticas y restos de fauna (Gorjanovic, 1906). Kaprina 5 (Caspari y Radovcic, 2006) muestra una gran robustez en contraste con otros cráneos de este yacimiento.
España
- Lezetxiki, Mondragón, Guipúzcoa. Primer molar y segundo premolar.
- Bolomor, Tavernes, Valencia. 5 dientes y fragmento de fémur.
- Tossal de la Font, Castellón. Dos restos poscraneales.
- Cova Negra de Bellús, Xátiva, Valencia. Nivel 33. Fragmento mandibular.
- Cova Foradá, Oliva, Valencia. Nivel 48. Hogar con restos de lepóridos y otra fauna allí consumida.
- Pinilla del Valle, Madrid. La Niña de Lozoya. Cuatro dientes de un neandertal de corta edad, con indicios de enterramiento intencionado (2011). 20 pares de astas de uros cerca de fogatas. En el mismo valle, Abrigo de Navalmaíllo y Cueva de la Buena Pinta.
- Cueva Corazón, Cañón de la Horadada (Mave, Palencia). Yacimiento musteriense. los restos indican diversas actividades ligadas al desollado, el desarticulado y el descarnado. Junto al aprovechamiento cárnico, la médula de los animales también sería explotada como muestran las marcas de percusión y los patrones de fracturación.
- Los Casares, Riba de Saelices, Guadalajara. Metacarpiano. Extraviado.
- Cueva de Valdegoba, Huérmeces del Cerro, Burgos. Mandíbula, varios dientes, falange de la mano, dos metatarsianos y otros, pertenecientes al menos a cinco individuos distintos.
- Carihuela, Granada. Carihuela-3. Hueso frontal de un niño, depositado por algún depredador.
- Cova Eirós, Triacastela, Lugo. Artefactos que fueron utilizados como armas de caza. Otros útiles se emplearon para despiece. Otra parte del utillaje sirvió para cortar y tallar madera, posiblemente para fabricar los mangos de las armas de caza. Otros artefactos se usaron para preparar pieles.
Datados en la glaciación del periodo OIS4 (74.000-59.000).
Francia
- Regourdou. Sepultura datada hace 70.000 años. Asociada a hallazgos líticos musterienses de tipo Ferrassie. Regourdou 1, Esqueleto parcial muy completo.
- La Quina. Fósiles humanos de al menos 27 individuos, con industria asociada. H5, Esqueleto incompleto de adulto. H9, Mandíbula de adulto. H18. Cráneo infantil.
- Baume Néron (Defleur, 1994). Dos dientes.
- Abri des Pechêurs. Seis dientes.
Hungría
- Subalyuk. Esqueleto poscraneal, mandíbula de adulto y cráneo infantil, con industria musteriense.
Chequia
- Ochoz. Mandíbula de adulto, molar y fragmentos de huesos poscraneales.
Italia
- Caverne delle Fate, Savona, Liguria. Frontal, occipital, zigomático, fragmentos de mandíbulas y dientes sueltos.
Próximo Oriente
- Shanidar. Ralph Solecki descubrió 9 esqueletos de neandertal entre 1953 y 1960 en la cueva de Shanidar (Irak). Su edad está comprendida entre los 70 y 40.000 años. Shanidar 4 fué enterrado con una ofrenda de flores. Shanidar 1 estaba parcialmente ciego y mutilado de un brazo; Algunos restos muestran fosa canina y el malar está ligeramente angulado.
- Kebara. Dorothy Garrod y Francis Turville-Petre excavaron la cueva en la década de 1930. En 1982, apareció aquí Moshe, un esqueleto neandertal muy completo. El hueso hioides está bien preservado y fue el primer hueso hioides neandertal encontrado.
Península Ibérica
- Covalejos, Cantabria. Dos dientes de muda.
- Cova Negra de Bellús, Xátiva, Valencia. Niveles 20-17. Restos de 7 individuos (4 niños, 1 adolescente y 1 adulto). Cráneo 4, radio y fragmento de peroné, con un hogar a techo.
Datados en el OIS 3c (59.000-45.000)
Francia
- La Ferrassie (Francia) dos esqueletos adultos de diferente sexo y unos cinco infantiles (tres niños, un recién nacido y un feto), que se empezaron a desenterrar por Capitan y Peyrony a principios de 1900. Los restos infantiles fueron publicados por Jean-Louis Heim al principio de los años 1980. Hay evidencias de enterramiento intencional. Se han datado en MIS 3, entre hace 54 ± 3 y 40 ± 2 ka (Guillaume Guérin et al, 2015).
- La Ferrassie 1. 1909. Esqueleto de adulto, cráneo completo con moño occipital, frente inclinada, grandes arcos superciliares y cara muy grande.
- La Ferrassie 2. 1910. Cráneo y esqueleto incompleto de hembra.
- La Ferrassie 3. 1912. Esqueleto infantil casi completo.
- La Ferrassie 4 y 4bis. 1912. Esqueleto infantil.
- La Ferrassie 5. 1920. Feto de siete meses.
- La Ferrassie 6. 1921. Infante de 3-5 años. El cráneo estaba separado más de 1 m.
- La Ferrassie 8. 1970. Esqueleto parcial de un infantil de unos dos años de edad, descubierto en 1970, aunque la mayor parte de los restos fueron recuperados en 1973 por el equipo de H. Delporte. En 2015 se ha publicado el redescubrimiento de restos adicionales en el Museo Nacional de Historia Natural de Francia y en el Musée d’Archéologie Nationale et Domaine National de Saint-Germain-en-Laye (Asier Gómez-Olivencia, Isabelle Crevecoeur y Antoine Balzeau, 2015).
- Cueva Malarnaud (Francia). Mandíbula de adulto.
- La Cotte de St. Helier en la isla de Jersey
Chequia
- Khulna (Chequia). Cráneos, mandíbula y poscraneales de al menos 11 individuos. Maxilar derecho, pragmento de parietal derecho y dientes aislados datados en 45,6 ka.
España
- Carihuela-2. Nivel VI.
- Sima de las Palomas, Cabezo Gordo, Torre Pacheco, Murcia. 89 restos, entre ellos diversos fragmentos craneales y mandibulares.
- Cova del Gegant de Sitges, Barcelona. Mandíbula.
- La Güelga, sector oriental del Macizo Asturiano.
- Boquete de Zafarraya, Alcaucín, Málaga. 3 mandíbulas, escápula, húmero, pubis, 3 fémures, tibia y diente lacteal de nueve individuos.
- Jarama VI.
- Valdegoba. 48,5 ka.
- El Sidrón, Piloña, Asturias. En contexto secundario en un sistema kárstico (de la Rasilla et al, 2014; Dataciones: Wood et al, 2013; de la Rasilla et al, 2013). Más de 2.400 fósiles (más de 200 dientes) de al menos 13 individuos: uno infantil (2-3 años) otro infantil casi completo (6-7 años, hijo de una de las mujeres del grupo y hermano del anterior) con molares de leche e incisivos definitivos incipientes), otro juvenil posiblemente masculino, dos de adolescentes varones, siete de jóvenes adultos (tres hembras, tres machos y uno indefinido). Los infantiles, sin dientes. Asociados a 415 artefactos líticos y 51 restos faunísticos. En 2013 se hallaron un fragmento craneal y un maxilar de adulto al que faltan los molares de uno de los lados.
Datados en el intervalo frío OIS 3b (45.000-35.800 años)
Francia
- Le Moustier.
- Le Moustier 1. Restos perdidos en un bombardeo de Berlín durante la segunda guerra mundial con expceción de un esqueleto de infantil masculino datado en 42,5 ka.
- Le Moustier 2. Bebé de unos 9 meses exhumado por Denis Peyrony en 1914. Reposaba en una fosa oval de unos 50 cm y 40 cm de profundidad. Identificado por Bruno Maureille en 1996 en los depósitos del Museo Nacional de Prehistoria de Francia. Datado en 40,9 ka.
- L’Hortus, Languedoc. Restos de unos veinte individuos datados en 40 ka.
- Abri des Pechêurs. Cuatro dientes.
- Bouffia Bonneval, La Chapelle-aux-Saints.
- LCS 1, Old Man, descubierto por Amédée y Jean Bouyssonie en 1908. tiene una antigüedad de 40 ka y una capacidad craneal de 1.620 cc. El esqueleto casi completo fué reconstruido por Marcellin Boule quien exageró los ragos anatómicos, popularizando el estereotípo de hombre mono. Fue enterrado intencionalmente (William Rendu et al, 2013)
- Dientes de al menos dos individuos juveniles y un adulto.
Italia
- Grotta di Fumane. Evidencias de obtención de plumas de aves para uso simbólico.
Chequia
- Dzeeraba Skala. Molar inferior. Datado con dudas en 40 ka años.
Bulgaria
- Bacho Kiro. Datado en 43 ka.
Croacia
- Vindija. Cráneos, mandíbula y poscraneales. Todos los restos esqueléticos provienen del estrato G, con la posible excepción del ramus mandibular Vi-11.52 que podría pertenecer a la capa I, más antigua. Estrato G3: Musteriense con algunos elementos del Paleolítico Superior. 43-38 ka. Estrato G1: Puntas de hueso Mladec y otros artefactos de hueso. Mezcla de tipos musteriense y del Paleolítico Superior. Punta de hueso Vi-3437 asociada a mandíbula Vi-207. Punta de piedra bifacial de material no-local parecida a las de la industria szeletiense. Interpretaciones:
- Mezcla de elementos musterienses y auriñacienses.
- Industria transicional.
- Material artificialmente mezclado (muy improbable).
Alemania
- Feldhofer, Neander. Pequeña cueva. En 1856 Johann Fuhlrott halló una bóveda craneal, huesos del fémur, parte de una pelvis, algunas costillas y huesos del brazo y hombros. El individuo al que pertenecían se rompió el antebrazo izquierdo en vida, dando como resultado que los huesos del brazo izquierdo son más cortos que el derecho.
- Königsaue, Alemania del Este. Se hallaron dos pequeños bulbos de brea de abedul, uno con una huella digital y otro con la impresión de un mango de madera. La brea de abedul es un adhesivo cuya obtención requiere un proceso técnico complejo.
Península Ibérica
- Carihuela-1. Nivel 5.
- Arrillor. Muela infantil.
- Axlor (Indusi, Dima, Vizcaya). Maxilar incompleto y fragmento de mandíbula con varios dientes, de un mismo individuo.
- El Castillo (Puente Viesgo, Cantabria). Más de 30 fósiles de unos doce individuos.
- Banyoles. Mandíbula de adulto avanzado, con dientes muy gastados.
- Gruta da Oliveira, Almonda. 4 fragmentos.
- Cova Negra de Bellús, Xátiva, Valencia. Niveles 13-11. 2 metatarsianos, fragmento de diáfasis femoral, parietal-2.
- Calvero de la Higuera (Madrid) Lozoya, dos incisivos, un canino y una muela de una menor de 2 años y medio (2011), datados en más de 40 ka. Hay indicios de que el cuerpo de la niña fue depositado intencionalmente.
Datados en el intervalo oscilante OIS 3a (35.800-27.000)
Se constata una reducción de la robustez facial, cambios en las extremidades superiores, probablemente relacionados con el uso de la tecnología y los adornos, y en la anatomía locomotora, que denotan una forma diferente de moverse por el paisaje. Todo ello indica un cambio hacia comportamientos modernos (Trinkaus, 2011).
Francia
- Arcy-sur-Cure. Datado en 33.800 años.
- Saint-Césaire, Charente-Maritimes. Esqueleto aplastado descubierto por Francois Leveque en 1979, en un entierro secundario. El cráneo está casi completo, faltándole la parte inferior. Se ha datado alrededor de 35 ka. Se encontraron herramientas chatelperronienses, pero existe cierto grado de incertidumbre con respecto a su asociación. Poco prognatismo medio-facial, cara plana y dientes reducidos. Además, aunque no prominente, muestra una pequeña barbilla, una característica que está presente sólo en los HAM. Los arcos superciliares son muy diferentes a los de los neandertales clásicos, la frente se inclina para atrás hacia la glabela y las órbitas de los ojos y la nariz son bastante grandes.
Bélgica
- Gruta de Spy d’Orneau. Cráneos, mandíbulas y poscraneales (dos esqueletos casi completos Spy 1 y Spy 2) descubiertos por Marcel de Puydt y Max Lohest en 1886. Datados entre 38-34 ka. El hallazgo sirvió para desacreditar la idea de que los neandertales sufrían una especie de disminución psíquica. Los cráneos tienen entre sí diferencias significativas, lo que indica el gran margen de variación que existía dentro de la misma población. Cabe la posibilidad de que estas diferencias fueran debidas a la presencia de nuevos genes en la respectiva población.
Península Ibérica
- Cueva de Gorham, Gibraltar. Forma parte de un conjunto de cuevas ocupadas por neandertales. Desde hace 125 ka, el clima fue similar al actual. Los neandertales explotaban un hábitat similar al actual de Doñana, con abundancia de presas pero también con depredadores como leones, linces, lobos y osos pardos. La costa se hallaba a unos 5 km. de la cueva. Los neandertales comían grandes mamíferos, con preferencia por la cabra montés, pero los restos más abundantes corresponden a conejos; también son muy abundantes los restos de aves. Comían también tortugas de tierra, piñones, lapas, mejillones, focas y delfines. Es de suponer que comiesen también frutas, raíces y larvas, que no dejaron rastro. Datados en 30-26 ka aparecen restos auriñacienses, lo que puede indicar la convivencia con sapiens. Las investigaciones han revelado una red de pequeñas cuevas utilizadas para las expediciones de caza ocasionalmente antes de regresar a la base.
- Devil’s Tower. Media docena de fósiles incluyendo un cráneo infantil icompleto y una mandíbula.
- Covalejos. Nivel 8.
- El Salt, Alcoi, Alicante. 6 dientes.
- Cueva de los Moros de Gabasa, Ribagorza, Huesca. Premolar, dos molares, metatarsiano, falange del pie, clavícula.
- Columbeira. Germen de primer molar.
- Figueira Brava, Sesimbra. Nivel C-2. Premolar superior.
- Salemas, Ponte de Lusa. Infantil: varios huesos y un maxilar con dientes y una muela de leche.
- Cova Negra de Bellús, Xátiva, Valencia. Niveles 9 y 8. Cráneos 3 y 5.
- Cova Foradá de Oliva. Valencia. Roca con forma de cabeza de oso cavernario hallada junto a restos humanos, con incisiones.
Últimos neandertales
En Zafarraya se ha hallado una mandíbula que muestra características neandertales clásicas, al carecer de barbilla, dientes delanteros relativamente grandes y una arcada dental grande. En 2011 Javier Baena et al comunicaron las dataciones de los hallazgos en la Cueva del Esquilleu, Liébana, Cantabria, España. Los restos más modernos ofrecen una datación de 20 ka. La tecnología lítica asociada corresponde al último musteriense. Según David Santamaría (2012) los neandertales se extinguieron en la Península Ibérica antes de hace 40 ka. RE Wood et al (2013) han publicado una nueva datación por el método del radiocarbono con protocolo de ultrafiltración, de los hallazgos de Cueva del Boquete de Zafarraya (Málaga) y Jarama VI (Sistema Central), dos de los últimos refugios neandertales. La nueva datación, de 45-50ka, de diferentes ejemplares, descarta la coincidencia con Homo sapiens en estos lugares y pone en duda anteriores dataciones que asignaban 36 ka a estos reductos en la Península Ibérica. El protoauriñaciense aparece en estos lugares con dataciones de 42 ka. También pone en duda la asignación a neandertales de herramientas y adornos en base a la datación.En Vindija (Croacia), se halló una mandíbula neandertal (Vi-207), datada en 33-32 ka (últimos neandertales, datados con seguridad).
- Wallace, John A., et al. «Did La Ferrassie I use his teeth as a tool? [and comments and reply].» Current’Anthropology (1975): 393-401.
- Smith, Fred H. The Neandertal remains from Krapina: a descriptive and comparative study. Diss. University of Michigan, 1976.
- Wolpoff, Milford H. «The Krapina dental remains.» American Journal of Physical Anthropology 50.1 (1979): 67-113.
- Puech, Pierre-François. «Tooth wear in La Ferrassie man.» Current’Anthropology 22.4
- (1981): 424-430.
- Wolpoff, Milford H., et al. «Upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia.» American Journal of Physical Anthropology 54.4 (1981): 499-545.
- Smith, Fred H., Donna C. Boyd, and Mirko Malez. «Additional upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia.» American Journal of Physical Anthropology 68.3 (1985): 375-383.
- Mellars, Paul. «Dating and correlating the French Mousterian (reply).» (1986): 113-114.
- Radovcic, J., et al. «The Krapina hominids: an illustrated catalog of skeletal collection.» Zagreb: Croatian Natural History Museum (1988).
- Mann, Alan E., Janet M. Monge, and Michelle Lampl. «Investigation into the relationship between perikymata counts and crown formation times.» American Journal of Physical Anthropology 86.2 (1991): 175-188.
- Stiner, Mary C. «The faunal remains from Grotta Guattari: a taphonomic perspective.» Current’Anthropology 32.2 (1991): 103-117.
- White, Tim D., et al. «The question of ritual cannibalism at Grotta Guattari [and comments and replies].» Current’Anthropology 32.2 (1991): 118-138.
- Simek, Jan F., and Fred H. Smith. «Chronological changes in stone tool assemblages from Krapina (Croatia).» Journal of human evolution 32.6 (1997): 561-575.
- Recheisl, W., and H. Seidler. «Volume of the Guattari 1 Neandertal Specimen (Monte Circeo).» Coll.’Antropol 23.2 (1999): 397-405.
- Ahern, James, et al. «New discoveries and interpretations of hominid fossils and artifacts from Vindija Cave, Croatia.» Journal of Human Evolution 46.1 (2004): 27-67.
Homo sapiens
De acuerdo con Lieberman (1998) el cráneo de un humano anatomicamente moderno (AMH) debe mostrar: una caja craneal globular, frente vertical, arcos supraciliares poco marcados, fosa canina y mentón pronunciado (Pampush y Daegling, 2016). Los especímenes más antiguos que exhiben algunas de estas carcterísticas han aparecido en Etiopía (Collard y Dembo, 2013). Los más importantes son Omo I, Omo II y Omo III, datados en 195 ka. En Kenia (Koobi Fora) se han encontrado los más antiguos seres cercanos a los humanos de aspecto moderno, con una antigüedad de 300-270 ka obtenida mediante series de uranio. La edad de 0,64 ma atribuida a los especímenes de Bodo llevaría más lejos aún el origen de nuestra especie si es que el “hombre de Bodo” debe considerarse ya como un ser humano de aspecto moderno. Y de otorgar esa condición al ejemplar de Danakil, entonces nos habríamos remontado a cifras que rondan el millón de años. Estas fechas quedarían ya lejos del rango en que se mueven los indicios proporcionados por los métodos moleculares. Hacia 90-80 ka aparecen especímenes con rasgos modernos en el Sur y en el Norte de África. Fuera de África, los restos más antiguos de humanos modernos proceden de Skhul y Qafzeh, en Israel.
En los orígenes de la especie (hace 240-200 ka, MIS 7), repetidas expansiones y contracciones poblacionales dieron lugar a la asimilación dentro de un único linaje de novedades biológicas y de comportamiento adquiridas localmente (Mirazon, 2016).Las poblaciones africanas son más diversas que las demás poblaciones humanas. Los fósiles humanos anatómicamente modernos más antiguos se recuperaron en Omo Kibish y Herto, Etiopía (datados en 195-160 ka) y los hallazgos genéticos, apoya la hipótesis de que África Oriental es el lugar de origen de la especie, hace unos 200 ka. Los fósiles son sustancialmente diferentes entre sí y respecto a los humanos más recientes. La diversidad que revelan se confirma con los fósiles datados en MIS 6 de Djebel Irhoud, Marruecos, y Ngaloba, Tanzania. Otras hipótesis apuntan a un origen espacial complejo (Schuster et al, 2010) o a un origen en África del Sur (Compton, 2011; Marean, 2015; Henn et al, 2011). Todos estos fósiles están asociados a herramientas clasificadas como Middle Stobe Age (MSA). Los fósiles de Herto se han recuperado de un contexto con elementos tanto de la MSA como Achelense. La datación de estos fósiles es anterior a la fecha del ancestro común más reciente de los humanos actuales, tanto si tenemos en cuenta el ADN-mt como el ADN-Y o el ADN nuclear.
Hasta 200 ka, se clasifican dentro de Homo helmei. El Hombre de Helme hace alusión al capitán R. E. de Helme, quien financió las excavaciones del zoólogo Thomas F. Dreyer en Florisbad, Bloemfontein, Sudáfrica en la primavera de 1932. Para Larry y Foley (2001) Homo helmei es un grupo monofilético africano, último antecesor de los neandertales y los humanos modernos. Los fósiles entre 200-130 ka, se clasifican como Homo sapiens idaltu. En 2011, Stringer y Harvati publicaron el estudio de un cráneo de H. sapiens hallado en Iwo Eleru (Nigeria) en 1965 y datado en 13 ka. Presenta las características que corresponden a los H. sapiens arcaicos, lo que sugiere su supervivencia hasta épocas muy recientes y su cruzamiento con los humanos modernos (Stojanowski, 2014).
Homo helmei (Dreyer, 1935)
- Eliye Springs KNM-ES 11693, cráneo encontrado el 1983 en Kenya. 0,3-0,2 ma. Características arcaicas: quilla sagital, cráneo aplanado, posición baja de la anchura máxima del cráneo, torus suborbital, marcada inserción de los músculos nucales, cara maciza y masiva. Características modernas: alta capacidad craneal, la forma del cráneo en vista superior y trasera, prominencias frontales presentes pero muy poco marcadas, pariental, escama temporal desarrollada, prolongación de la parte superior del occipital, ausencia de torus occipital transversal. Rasgos patológicos sugieren anemia crónica (Bräuer et al, 2003).
- Koobi Fora. ER-3884. Cráneo datado en 0,27 ma.
- Florisbad, encontrado en Orange, Africa del Sur en 1932. Datado en 0,259 ma. Cráneo de un individuo devorado por un carnívoro de gran tamaño. Cráneo ancho, frontal algo deprimido con relieve supraorbital muy pronunciado, aunque se atenúa lateralmente y muestra una depresión en el plano sagital. Rostro ancho y robusto. Asociado a arqueología posiblemente MSA temprana.
- Cave of Hearths (Cueva de los Hogares, África del Sur). Mandíbula. 0,2 ma. Asociado a arqueología achelense o MSA temprana.
- Littorina Cave, Sidi Abderrahman (Casablanca, Marruecos). Hemimandíbula asociada a industria achelense. Más de 0,2 ma.
- Qebibat. Mandíbula. 0,25 Ma.
- Omo I, II y III, encontrados en 1967 en la Formación Kibish, Etiopía, por Richard Leakey. (Brown y Fuller 2008 ; McDougall, Brown, y Fleagle 2005 ; Shea 2008). Asociados a industrias de la MSA. Brauer (1991) propuso clasificar Omo I como anatómicamente moderno y Omo II como sapiens arcaico reciente. Para Tobias son contemporáneos y dan idea de la varibilidad de la población.
- Omo I. Ftagmento de cráneo de un joven varón, incluyendo fragmentos de la cara y parte del esqueleto postcraeal. Reconstruido por Day y Stringer (1982). Morfología general muy moderna. Frontal recto, bóveda elevada y regularmente redondeada. El occipital presenta una parte superior mucho más larga que la parte inferior. Costados divergentes hacia arriba. Superestructuras óseas poco desarrolladas. Mentón típico. >1.400 cc. Costillas largas. 70 kg. Datado en hace 195 ka por 40Ar/39Ar (Day y Stringer, 1991; McDougall, Brown y Fleagle 2005; Pearson et al, 2008; Voisin, 2008).
- Omo II. Cráneo de un adulto varón hallado a 2,5 km del anterior. Torus supraorbital pequeño. Presenta varios rasgos arcaicos como la gran angulación del occipital o el grosor del hueso timpánico. 1.435 cc. Para Day y Stringer, mientras que Omo I es Homo sapiens strictu senso, Omo II no lo es. Datado en hace 195 ka, por correlación estratigráfica con Omo I. Feibel, 2008; Rightmire, 2009.
- Omo III. Calvaria fragmentada de un adulto de sexo incierto.
Homo sapiens idaltu (White et al, 2003)
Presentan un prognatismo nasal más reducido, mayor capacidad craneal y una frente más elevada y convexa.
- Jebel Irhoud (Marruecos 1961). Restos de 5 individuos entre 190-90 ka. No ha podido establecerse una continuidad entre estos restos y los restos aterienses posteriores. Paleoentorno (Geraads et al, 2013).
- Jebel Irhoud I es el cráneo más completo. Se puede observar un neurocráneo primitivo con una gran capacidad craneal de 1.305 cc y cara con una morfología muy moderna.
- Jebel Irhoud 2 (Marruecos 1961). Restos datados en 160 ka atribuidos al musteriense (estadio isotópico 5, Hublin 1991). Cráneo y bóveda craneal de adultos y mandíbula, fragmento de húmero e ilion de niños. Arquitectura craneal moderna: bóveda elevada, frontal recto, región occipital bastante redondeada, paredes que divergen hacia arriba con protuberancias parietales muy desarrolladas. Se presentan también rasgos arcaicos: relieve supraorbital robusto y continuo.
- Miembro Herto de la Formaciñon Bouri (Middle Awash, 2003). Tres cráneos (dos hombres y un niño) con marcas de corte que evidencian un canibalismo ritual o prácticas simbólicas ligadas a ritos mortuorios. Datados en 160-154 ka por 40Ar/39Ar. Tim White et al, los clasificaron como homo sapiens idaltu. (Clark et al, 2003. White et al, 2003). Muchos fragmentos craneales muestran marcas de instrumentos líticos, lo que se interpreta como evidencia de algún tipo de prácticas mortuorias descartándose, por su patrón de distribución y su morfología, el consumo.
- BOU-VP-16/1. Descubierto en 1997. Cráneo adulto prácticamente íntegro. Mezcla de rasgos primitivos y modernos. Frente vertical y neurocráneo globular, dentro del rango humano moderno. La mayor anchura craneal se produce en la parte superior de los parietales, como en los humanos actuales. 1.450 cc.
- BOU-VP-16/2- Restos craneales de un adulto.
- BOU-VP-16/5. Cráneo juvenil incompleto (180 fragmentos).
- Presentan un mosaico de rasgos
- Primitivos: Hueso occipital muy angulado, con un torus superior muy marcado, torus supraorbitario muy marcado.
- Derivados: Volumen encefálico elevado, bóveda craneal alta y globosa, máxima altura del cráneo en los huesos parietales, hueso frontal alto, ausencia de prognatismo y placas infraorbitarias orientadas coronalmente mostrando una fosa canina.
- Según White et al (2003) estos especímenes son intermedios entre Broken Hill y los hallazgos datados en unos 100 ka.
- Singa (Sudán). >133 ka por ESR y U/Th. Cráneo bastante completo y patológico (Grün y Stringer, 1991; McDermott et al, 1996; Spoor et al, 1998; Stringer, 2003; Bräuer et al, 2004; Trinkaus, 2005; Bastir et al, 2008; Schwartz y Tattersall, 2010).
Homo sapiens anatómicamente modernos (130-100 ka).
- LH 18 (Laetoli Hominid 18, encontrado el 1976 en Ngaloba, Laetoli, Tanzania. >0,12 ma. Frontal oblicuo y relativamente aplastado, con relieve supraorbital poderoso y continuo. Las paredes craneales tienen un considerable grosor. La región occipital es regularmente convexa y el rostro no voluminoso (Day, Leakey y Magori, 1980).
- Mumba XXI (Tanzania 1977). Varios molares. Bräuer y Mehlman (1988), Mehlman (1989).
- Equus Cave (África del Sur 1980)
- Die Kelders Cave (África del Sur 1980)
- Border Cave (África del Sur, 1940, 42, 76).
- Cráneo y varios poscraneales de adulto masculino.
- Mandíbula, otros fragmentos de cráneo, mandíbula y poscraneales de un niño.
- Resto mandibular.
- Blind River, África del Sur. 118 ka. Asociado a artefactos.
Según Ilan Gronau (2011), los San se separaron del resto de las poblaciones africanas hace aproximadamente 130 ka.
Norte de África
Hace 130 ka el Sahara era una zona de lagos y riveras. En este tiempo y lugar surgen las primeras formas inequívocas de comportamiento moderno conocidas hasta el momento. Los yacimientos aterienses se extienden de Marruecos a Libia con una datación entre 145-70 ka. En la Cueva de los Contrabandistas (Costa atlántica, Marruecos) se halló en 2011 el cráneo de Bouchra, de un individuo de unos 6-8 años de edad, datado en 108 ka. Tiene unos dientes grandes y esta característica lo relaciona con los primeros H. sapiens que abandonaron África. Para Hublin es posible que poblaciones aterienses se estableciesen en el este de África para desde allí iniciar el out of Africa hace 60 ka. Hublin et al (2012) analizaron más de 50 dientes hallados en yacimientos aterienses encontrando un parecido sorprendente con los del cráneo y mandíbula de Pestera cu Oase, correspondiente a los europeos más tempranos. El cráneo de Nazlet Khater (Egipto), datado en 40 ka liga también a los aterienses con los europeos tempranos (Hublin y Klein, 2011). Por otra parte, puede haber una relación con las poblaciones de Oriente Medio: el cráneo Dar-es-Soltan 5 (Rabat, Marruecos), datado en 80 ka es muy similar al de Qafzeh, datado en 100 ka, con torus supraorbital pero con características modernas, como una dentición reducida en tamaño. También en Dar-es-Soultane se halló una mandíbula (Ferenbach, 1976; Hublin, 1993, 2000; Cartmill y Smith, 2009). De datación similar son el cráneo y mandíbula de Témara (Rabat, Marruecos) (Ferenbach, 1976; Hublin, 1993; Cartmill y Smith, 2009). Témara 2 es un craneo fragmentario con un occipital redondeado y carece de torus supraorbital. La mandíbula es moderna.
- Grün R, Stringer CB (1991) Electron spin resonance dating and the evolution of modern humans. Archaeometry 33(2), 153-199.
- Hublin J-J (1992) Recent human evolution in northwestern Africa. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences 337(1280), 185-191.
- Clark JD, et al (2003) Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423(6941), 747-752.
- White TD, et al (2003) Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423(6941), 742-747.
- Smith TM, et al (2007) Earliest evidence of modern human life history in North African early Homo sapiens. PNAS 104(15), 6128-6133.
- Butzer KW (1978) Sediment stratigraphy of the Middle Stone Age sequence at Klasies River Mouth, Tsitsikama Coast, South Africa. The South African Archaeological Bulletin 33(128), 141-151.
- Rightmire GP, Hilary JD (1991) Comparative studies of Late Pleistocene human remains from Klasies River Mouth, South Africa. J Hum Evol 20(2), 131-156.
- Lam YM, Osbjorn MP, Smith CM (1996) Chin morphology and sexual dimorphism in the fossil hominid mandible sample from Klasies River Mouth. Am J Phys Anthropol 100(4), 545-557.
- Grine FE, et al (1998) Additional human fossils from Klasies River Mouth, South Africa. J Hum Evol 35(1), 95-107.
