La dispersión y diversificación de las poblaciones humanas modernas responde a múltiples factures, incluyendo selección natural, aislamiento reproductor, deriva genética, efecto fundador, etc. La aparición de los caracteres fenotípicos raciales (de las poblaciones de humanos modernos) responde a procesos de adaptación ambiental por selección natural en función de variables geográficas y climáticas que se distribuyen geográficamente en forma de clinas (gradientes), por lo que la frecuencia y la expresión fenotípica de dichos caracteres también se distribuye formado un gradiente continuo de variabilidad, sin que existan grupos discretos y aislados desde el punto de vista biológico. Además, las barreras culturales determinan una diversificación poblacional que generalmente es superior a las diferencias biológicas. Aun así, las sociedades humanas tradicionales presentan una gran variabilidad morfológica que es posible caracterizar y que deriva de su historia evolutiva y su dinámica poblacional particulares. Los procesos evolutivos y de diversificación genética que han actuado en cada momento y en cada zona geográfica, y su velocidad, han variado enormemente. El análisis de la diversidad poblacional humana actual permite caracterizar qué procesos han actuado en cada momento, con el fin de entender el origen de nuestra diversidad biológica.
Diversificación de Homo sapiens moderno
Los Homo sapiens modernos más antiguos data de hace unos 200.000 años, con una transición morfológica en diversas zonas a partir de Homo erectus. Esta transición también es cultural: la técnica Levallois desapareció con las nuevas técnicas de tallado (Auriñaciense o modo 4) que llegaron a Europa hace 40.000 años. A partir de 0,3 ma ya no hay Homo erectus. Los restos más recientes son los de Choukoutien (China). La transición no se hizo uniformemente ni a la misma velocidad en las diferentes áreas geográficas. Homo erectus se mezcla en unos casos y se extingue en otros. En Java sus caracteres persistieron hasta época muy reciente por la insularidad. También hay alguna evidencia de mezcla con Homo sapiens arcaicos (bóvedas de Ngandong o de Solo). Se han planteado diversas hipótesis sobre el origen de los humanos modernos.

- El jardín del Edén o El Arca de Noé: visión extrema de substitución de las poblaciones arcaicas por los humanos modernos. Se suele llamar hipótesis del reemplazamiento, liderada por C.B. Stringer principalmente. Los genetistas moleculares la han apoyado tradicionalmente, basándose tanto en los datos como en la visión de expansiones démicas tras la adquisición de la agricultura. La limitación de la Genética Molecular en este tema es que no detecta los alelos perdidos (o variaciones del mtDNA) de las poblaciones antiguas que no han dejado descendencia.
- La hipótesis Afro-Europea: del alemán G. Bräuer, versión atenuada de la anterior. Propone un origen Africano que tempranamente entró en Europa, donde por aislamiento dio origen a los Neandertales (H. sapiens neanderthalensis). Coincide con la anterior en que grupos de poblaciones modernas substituyeron progresivamente a las menos avanzadas tecnológicamente. Difiere en que acepta mezclas locales en los extremos de los continentes (Europa, Japón, Java y Australia).
- Hipótesis Multiregional: opuesta al Jardín del Eden. La propuso en los años 40 F. Weidenreich. Los grupos raciales actuales habrían evolucionado localmente, limitados por la geografía, desde H. erectus. Tiene pocos seguidores, uno de ellos era Carleton Coon (su libro Las Razas Humanas, Ed. Guadarrama, fue traducido por A. Valls), pero esta hipótesis es el origen de la siguiente.
- Hipótesis de hibridación por flujo génico: versión atenuada de la anterior, propuesta por M. Wolpoff, D. Frayer y A. Thorn. Subraya la importancia del flujo génico y del paralelismo adaptativo y cultural. Wolpoff suprime H. erectus y lo incluye en Homo sapiens, que habría existido desde hace 1.8 ma. Sin embargo, no explica adecuadamente la 1) enorme variación intra-grupal de la especie, ni 2) la variación clinal de las poblaciones modernas actuales, producto de la evolución local. Según esta hipótesis los europeos actuales vendrían de los Neandertales.
Homo sapiens arcaico
Se conocen pocos especímenes transicionales entre Homo erectus y Homo sapiens, como Bodo (Etiopía) de 600.000 años, o como Salé, Rabat y Casablanca (Sidi-Abder-Rhaman) de unos 400.000 años (aunque esta datación está muy discutida). Son diversos los candidatos a primeros Homo sapiens arcaicos:
- África: Bodo (Etiopía), Ndutu (Tanzania), Kabwe (antes conocido como Broken Hill, Zambia), Hopefield (antiguo Saldanha, de Sudáfrica). Las datación de estos fósiles varía entre 400.000 y 200.000 años. Stringer clasifica Bodo y Kabwe como Homo heidelbergensis africano, mientras que otros los consideran H. rhodesiensis o simplemente como H. sapiens arcaico.
- Europa: Aragó (sur de Francia), Ceprano (Italia) y Verteszollos (Hungría), de hasta 450.000 años. (Petralona datado en 0.2 ma). En realidad algunas de estas formas serían antecesoras de los Neandertales y se clasifican como H. heidelbergensis europeos.
- Java: 13 bóvedas de Ngandong o de Solo, probable mezcla con Homo erectus. De unos 150.000 años.
- China: Dali, de 0.3-0.2 ma, Ying-Kou (esqueleto), Mapa (o Maba) y Chang-Yang. Muestran algunos caracteres derivados de los humanos modernos y otros arcaicos.

Homo neanderthalensis
Los primeros poblamientos de Europa probablemente no fueron permanentes. Las más antiguas ocupaciones están representadas por las Industrias de Barranco León y Fuente Nueva (Orce), de hace 1.000.000 años aproximadamente y los materiales de Atapuerca (Sima del Elefante, 1.1 ma; Gran Dolina, 0.7 ma) y Boxgrove (sur de Inglaterra, 0.5 ma). La presencia permanente de los huma-nos antes de 500.000 años en Europa es escasa. Según la hipótesis Afro-Europea del origen de Homo sapiens, habrían sido incursiones de Homo erectus. Hoy se descarta la antigua distinción entre Ante-Neandertales (sin rasgos de los posteriores Neandertales) y Pre-Neandertales (con rasgos de los posteriores Neandertales) de M.A. Lumley.
Los Neandertales sensu stricto serían los pobladores de Europa y Próximo Oriente entre hace 80.000 y 35.000 años. Los Pre-Neandertales serían todos los fósiles europeos que les precedieron, desde hace 450.000 hasta hace 80.000 años. La mayoría de estos materiales actualmente se clasifican como H. heidelbergensis. Para algunos autores, la mandíbula de Mauer y los restos de Bilzingsleben, ambos en Alemania, podrían ser formas transicionales de Homo erectus. Aragó 21 tiene ya la pneumatización de la cara que caracterizará a los Neandertales, pero se ha sugerido que su coxal tiene rasgos de Homo erectus. El occipital de Verteszollos muestra aún una gran área de inserción nucal propia de Homo erectus. Sin embargo, en estos fósiles se advierte ya un proceso de neandertalización gradual, evidente en toda la muestra euro-pea, que culmina hace 120.000 años en los especímenes de Saccopastore (Italia) y Tabun (Próximo Oriente), a donde emigraron grupos de Neandertales, originando así las poblaciones extraeuropeas de Neandertales. Otros restos atribuibles a los Preneandertales son Steinheim (Alemania), Swanscombe (Inglaterra), Petralona (Gre-cia), Eringsdorf (Alemania) y Saccopastore, que son los más recientes (pre-Wurm, anteriores a 120.000 años pero considerados por algunos como los primeros Neandertales). Otrosd restos más enigmáticos son Montmaurin y Banyoles. La datación del travertino de la mandíbula de Banyoles es de 40.000 años, lo que no coincide ni con su morfología ni con la uniformidad de los Neandertales en ese momento en Europa. Los Neandertales estuvieron aislados en Europa desde hace 400.000 años hasta hace 80.000 a 35.000 años.
Homo sapiens moderno
Se distinguen por dos caracteres: 1) gracilidad ósea (respecto a los anteriores) que en parte se atribuye a los adelantos técnicos que alejarían al cazador de la presa (boleadoras, lanzas, etc.; otros avances como el arco y la flecha, propulsor o boomerang, parecen ser posteriores) y 2) la aparición de eminencia mentoniana por reducción del tamaño de la dentición anterior (se reducen las raíces y es posible un refuerzo óseo ex-terno). Los fósiles más antiguos retienen aún rasgos arcaicos.
- Africa del este: Omo II, Omo III, Ngaloba (Laetoli 18), ES116963 Florisbad, Diré-Dawa. Los posteriores son indistinguibles de los humanos actuales.
- Africa del sur: Border Cave, Klasies River Mouth (datados en 0.12 ma).
- Próximo Oriente: Qafzeh 6 y 9, Skhuk 5 (90.000 años) muestran ciertos rasgos arcaicos (probable flujo génico). Su cultura fue Musteriense (de los Neandertales), pero son claramente humanos modernos por cráneo y por mantener proporciones «cálidas» en las extremidades.
Tradicionalmente se ha considerado que los Neandertales fueron substituidos por los Cromañones en Europa, que habrían llegado a Europa hace 40.000 años portando una tecnología más avanzada (el Auriñaciense) que la Musteriense de los Neandertales.
Los rasgos cromañoides se dan en todos los primeros Homo sapiens modernos extraeuropeos de cualquier área, por lo que no tienen valor poblacional; las poblaciones del Paleolítico Superior europeo (entre 40.000 y 10.000 años) substituyeron a los Neandertales, tanto en P. Oriente como en Europa; el origen de la actual población europea procede en su mayor parte de la expansión geográfica y démica del Neolítico procedente del Próximo Oriente hace unos 6000 años. Por tanto, las poblaciones del Paleolítico Superior en Europa no han contribuido significativamente a la actual. En China las poblaciones actuales presentan incisivos en pala, rasgo que ya se da en Homo erectus pekinensis de Choukoutien hace 0,3 ma. Aunque esto se ha utilizado como argumento a favor de hipótesis Multiregional, se podría explicar también por mezcla con Homo sapiens en China. Australia y Melanesia se poblaron hace unos 40.000 años. En el interior de Australia hay fechas de 38.000 años atribuidas a pinturas rupestres en cuevas, así como instrumentos equivalentes a los del Paleolítico Superior europeo. Los Aborígenes Australianos presentan rasgos dentales del Pleistoceno Medio (gran dentición anterior), gran robustez craneal, pero gracilidad corporal, y uso de útiles a gran distancia (propulsor y boomerang).
La transición de H. sapiens arcaico hacia H. sapiens moderno se produjo hace más de 200.000 años en África. En Europa coexistieron con los Neandertales (hasta hace 35.000 años). Hace 200.000 años aparecen formas transicionales que podemos asociar claramente a Homo sapiens sapiens.
África
En África se encuentran las formas de H. sapiens más antiguas, aunque es difícil diferenciar algunas de sus características de las de los arcaicos. Lo más antiguo Omo (al sur de Etiopía), quizá con continuidad desde formas arcaicas a modernas (Omo 1 tendría unos 200.000 años), seguido en África del este por Herto (165.000 años).
En Sudáfrica Florisbad y Klasies River Mouth podrían ser antiguo, con caracteres similares a los de los Aborígenes Australianos actuales), mientras que Bor-der Cave 1 es claramente moderno, aunque sus dataciones son controvertidas. Las formas africanas próximas a los humanos modernos se habrían expandido hacia el norte hace en torno a 100.000 años, cruzando el Sáhara hacia Marruecos y Próximo Oriente en una fase climática húmeda. En el sur de África habría habido una ocupación continuada hasta los actuales pueblos Khoisan (Bush-Hotentoes). Algunos cráneos de 35.000 años de Ciudad del Cabo y Zimbabwe pueden ser clasificados como Bosquimanos. La relativa robustez de estos grupos indicaría aislamiento. Así, al norte habría existido un grupo de negroides robustos (Semliki River, frontera de Zaire y Huganda) hace 25.000 años, distintos a los bosquimanos. Excepto en el desierto del Kalahari, los bosquimanos han desaparecido, en parte desplazados por los Bantu (agricultores y ganaderos nómadas del Norte) y los europeos. La expansión bantú se habría producido durante el último milenio desde el oeste de África llevando la ganadería y la agricultura al Este y Sur de África.
En el norte de África, Jebel Irhoud (Marruecos, 190.000 años) es similar a los Neandertales. Singa (en el Nilo Azul en Sudán, 133.000 años) no presenta caracteres neandertales. Dar es-Soltan (Marruecos, 80.000 años) no sería neandertal según Hublin. Haua Fteah (libia, 47.000 años) sería arcaico pero no neandertal. Para Hublin todos los restos del norte de África serían similares a las formas arcaicas de sapiens de Próximo Oriente (Skhül-Qafza).
Próximo Oriente
En Próximo Oriente, Zuttiyeh (Galilea, 250-350.000 años) es un fósil discutido que no parece asociarse a neandertal ni a humano moderno. Wolpoff ve similitudes con H. erectus de Zhoukoudian. Qafza 100.000 BP, con unos 20 individuos, y
Skhül 82.000 BP (Mt. Carmel, Israel), con al menos 10 individuos, son claramente humanos modernos. Sin embargo, Tabun (muy cerca de Skhül) es Neandertal y su datación es de 100.000 años, lo que indica una dinámica compleja de ocupación. Qafza 6 y 9 son más modernos ,aunque Skhül 5 es más neandertal y con algunos caracteres arcaicos (torus supraorbitario). Los modernos de Próximo Oriente habrían sido sustituidos por Neandertales hace unos 80.000 años. Amud y Kebara (50.000 BP) son neandertales. Aparecen durante el último periodo glacial y los modernos desaparecen del Levante. Para Trinkaus, sin embargo, Shanidar es una muestra de continuidad preneandertal-neandertal. En cualquier caso, la discontinuidad poblacional no se observa en las industrias que son Musterienses tanto en modernos como en neandertales.
Australia
Australia probablemente fue ocupada por humanos modernos desde África por Arabia y el Sudeste Asiático. Homo erectus no pudo cruzar la línea de Wallace (separa mamíferos de marsupiales de Australia) pero si pudieron los modernos hace unos 75.000 años (Arnhem en Australia. Homo erectus de Solo (Java) está datado en sólo 53.000 BP. Los yacimientos con restos humanos más antiguos datan de hace 55.000 BP pero los restos humanos en Australia más antiguos son de Lake Mungo (26-30.000 BP), con dos cráneos bastante gráciles; y también en Keilor (13.000 BP), un cráneo grácil. Ambos difieren de los Aborígenes Australianos actuales. Sin embargo, en Kow Swamp (10.000 BP) hay caracteres arcaicos muy robustos en la región craneal (no en el postcraneal); también es robusto un individuo de Lake Nitchie (al norte de Sidney, 6.000 BP, antes de la llegada del Dingo hace 5.000 años que traen nuevos pobladores. Los pobladores de Tasmania eran tan gráciles como los actuales australianos y llegaron hace unos 30.000 años quedando aislados hace 8.000 años por subida del nivel del mar. Esto indicaría que los primeros humanos modernos en Australia eran gráciles.
Grupos dispersos del Sudeste Asiático podrían ser descendientes de las migraciones a Australia hace unos 75.000 años (Papúes negritos del altiplano de Nueva Guinea, aunque podrían ser una adaptación al altiplano), que coincide con una erupción volcánica en Toba (Sumatra) hace 73.500 años. Las formas gráciles se detectan en Niah Cave (Sarawak, Borneo, quizá 40.000 BP), Tabon Cave (Palawan, Fili-pinas, 23.000 BP) y Waja (Java). Estos grupos llegaron antes de que formas asiáticas llegaran. Finalmente, también está clara la llegada hace unos 5.000 años de nuevas migraciones Austronésicas, que ocuparon la costa de Nueva Guinea.
Europa
En Europa la aparición de los humanos modernos supone la desaparición de los Neandertales. El Musteriense es sustituido por el Augriñacience y una gran riqueza de ejecución artística y de materias primas (hueso, conchas, dientes, lítico). Aparece el arte rupestre (los Neandertales sólo usaron el ocre quizá para pintarse los cuerpos, en los enterramientos). La transición no está clara, pero los europeos del Paleolítico Superior difieren de los neandertales que les preceden, y también de los europeos modernos (tienen dientes más grandes y cráneos más robustos). Su entrada en Europa no está clara. Las zonas frías de Europa eran difíciles de ocupar. Llegan hace 35.000 años y reemplazan a los Neandertales. En Europa Central aparecen restos de modernos con caracteres de neandertal (Vindija, Mladec, Predmostí) y otros ya con rasgos modernos (Chancelade, Abrie Pataud, Cro-Magnon, Les Eyzies). Otros ya con rasgos modernos son Chancelade, Abrie Pataud (Dordogna, Francia), Cro-Magnon (40.000 BP), Les Eyzies (Dordogne, Francia). Posteriormente se encuentran restos en torno a los 10.000 BP en Muge (Portugal) y en Bretaña África. Estas formas recientes son ya plenamente europeas (caucasoides).
Asia
En Asia pueblos se diferencias varios grupos humanos modernos:
Pueblos sundadontos y mongoloides: los Negritos son cazadores de baja estatura y mucha pilosidad de las islas Filipinas, desde Luzon a Palawan (incluso Taiwan). Grupos similares hay en las montañas de la Península Malaya (relacionados con grupos del Este de la India y Ceylán) y también los pigmeos de las Islas de Andaman (al Sur de Burma en el Océano Índico). Los andamaneses se asemejan a los africanos en la pigmentación de la piel, la pilosidad y en la esteatopigia, pero no se parecen estrictamente a los pigmeos africanos. En el Sudeste Asiático (Indonesia y Filipinas) se observan grupos con pilosidad y se podría aceptar mezcla con Negritos o Melanesios hoy extinguidos. En la India también se observan restos de este estrato antiguo pero muy mezclado con los caucasoides (hoy pigmentados) que entraron por el noroeste. Los caucasoides también llegaron a Asia Central (Iran, Ayrans, caucasoides con lengua indoeuropea). Los mongoloides derivarían de pueblos nómadas del este enfrentados tanto a los caucasoides del oeste como a los pueblos chinos emergentes. Las formas iniciales de mongoloides ocuparían el sur (Paleomongólidos). Los mongólidos del norte derivarían de la intensificación de los caracteres (cara plana, pliegue parpebral) hacia el norte (Coreanos, China, Japón y Este de Siberia), probablemente como adaptación progresiva al frío. Estudios dentarios (C Turner) definen el patrón Sundadonto por las presencia de 30 caracteres dentarios complejos (como dientes en pala, sexta cúspide en molares inferiores, etc.), típico delas formas paleo-mongólidas. Estos caracteres dentarios no están presentes en los australianos, africanos ni europeos. Los neo-mongólidos, que son Sinodontos, tienen un patrón aún más complejo. Son, por tanto, estadios del mismo desarrollo progresivo (unos descienden de otros). Restos de sundadontos se detectan en los aborígenes de Taiwan (los Bunun son particularmente cortos y pigmentados), en los Jomon (antiguos pobladores de las islas del Japón), cuyos descendientes son los Ainú (tienen considerable pilosidad). Los Ainú llegaron a Japón caminando (no había islas) hace 10.000 años y en toda la zona habría poblaciones Negrito y Australoides. Las poblaciones sundadontas representarían el tercer centro independiente de invención de la agricultura y habrían desplazado al estrato cazados anterior.
Pueblos sinodontos: las poblaciones neo-mongólidas se distinguen claramente del patrón paleo-mongólido y australo-melanesio. Los japoneses actuales llegaron desde Korea justo antes de la Era Cristiana, portando el cultivo del arroz y sustituyendo a los Jomones indígenas y relegando a los Ainú. Los Koreanos también llegaron tardíamente (son neo-mongólidos). Los Chinos en cambio se remontan a hace más de 4.000 años y se consolidan con la dinastía Han (206 BC) expandiéndose hacia el Sur progresivamente. Los chinos del Norte se habrían asentado hace ya 7.000 años. Las evidencias más antiguas son escasas.
Habría dos localidades significativas con humanos moderno: Zhoukoudian (Upper Cave 101 ‘Old man’ y varios cráneos más, 25.000 BP, difícil de clasificar como mongoloide; y Upper Cave 102 más mongoloide en apariencia) y Liujiang (Sur de China, 67.000 BP, es moderno en todo excepto en un torus orbitario bastante robusto: sería un proto-mongoloide tipo sundadonto, coetáneo con los neandertales en Europa y con los primeros pobladores australianos).
Oceanía
En Oceanía las poblaciones de lenguas Austronésicas son relativamente recientes. Se encuentran en las costas de Nueva Guinea y Melanesia. Las lenguas austronésicas difieren del la de los Papúes de Nueva Guinea. Estos podrían haber llegado hace unos 40.000 años y serían responsables del desarrollo de sistemas de irrigación para la agricultura datados en 9.000 BP, en el altiplano de Nueva Guinea, y domesticación del cerdo hace 8.000 años. Algunas lenguas Papúes sobreviven en las Islas Salomon y habrían llegado incluso hasta Nueva Caledonia. Los austronesios llegaron después y ocuparos las costas de Nueva Guinea (no el altiplano ya ocupado por poblaciones que quedaron aisladas). Los polinesios, en cambio, llegaron hasta Nueva Zelanda, Pascua y Hawaii. Todos hablan lenguas austronésicas pero difieren algo morfológicamente (son más robustos en el tronco) probablemente por acción de la selección natural (vientos fríos del Pacífico a pesar de proceder de zonas ecuatoriales). Los polinesios podrían proceder de cualquier punto entre Taiwan y el Este de Indonesia, hace alrededor de 2.000 años (cultura Lapita ‘pottery’ asociada). Hace 1.500 años habrían llegado a Fiji, Tonga y Samoa procedentes de Salomon. Entre 400 AC y 1.200 AC llegarían a Pascua, Hawaii y Nueva Zelanda. No habrían llegado más lejos quizá por encontrar zonas ya ocupadas.
América
Los nativos de América cruzaron la región de Bering desde el Norte de Asia, lo que es actualmente territorio neo-mongólido. Sin embargo los amerindios actuales no son neo-mongólidos (aunque sí tienen caracteres asiáticos) y se han clasificado como marcadamente sinodontos (carácter también presente en Chukutien UP 101). En cambio los Eskimales americanos son muy parecidos a los Chukchi (esquimales Siberianos). Habrían llegado hace unos pocos miles de años. Turner (dientes), Zegura (genes nucleares) y Greenberg (lingüista) proponen un esquema de tres migraciones desde el Noreste de Asia: Los Esquimo-Aleuts serían los últimos en llegar, los NaDene (principalmente los Atabascanos del Noroeste y Navajo y Apache del Sudoeste) serían anteriores (pocos miles de años antes, con lengua NaDene de origen Sino-Tibetano) y los más antiguos serían los Ameríndios (lenguas ameríndias) con cultura paleoindia anteriores a unos 10.000 años.
Este panorama es demasiado sencillo y actualmente está muy discutido, ya que la disimilitud morfológica entre ameríndios y neo-mongólidos es bastante marcada (en caracteres faciales y corporales). Además, los estudios de Marta Lahr en Fueguinos sugieren similitudes entre la robustez de algunos fueguinos y formas prehistóricas del Norte de África (Afalou) y en cierta forma también Australia. Podrían representar una primera migración marcada por caracteres robustos similares a los observados en Upper Cave 101 en Zhoukoudian, pero actualmente se considera convergencia entre poblaciones que retienen rasgos arcaicos.
La cultura Clovis está documentada en Norteamérica hace 12.000 años. La entrada por Beringia habría sido posible con anterioridad a través de praderas que unirían ambos continentes (nivel del mas bajo durante el Pleistoceno final). Hay dataciones fiables de 20.000 BP en Pensylvania y 30.000 en Brasil (pre-Clovis), pero son yacimientos demasiado aislados. Datos de DNA mitocondrial de Fueguinos parecen sugerir que algunos linajes fueguinos difieren de los de los ameríndios, apoyando la hipótesis de una migración anterior.
La diversificación de las poblaciones humanas actuales
Variabilidad poblacional en Africa
 Existen grupos pigmentados en África, Asia (India) y en Oceanía (Melanesios). Aunque se ha postulado un mismo origen para todos ellos, podría tratarse de convergencia por adaptación climática (Andamaneses). El norte de África está ocupado por grupos de influencia caucasoide y zonas de contacto en el Sáhara (saharianos, tuareg, tebú y etiópidos). Al Sur están las poblaciones negroides. El África subsahariana comprende tanto zonas de sabana, desierto y bosque tropical. Los Pigmeos y los Khoisánidos o Káfridos (Hotentotes) no pueden considerarse poblaciones negras melanodermas. Las poblaciones negras son los Sudánidos, Nilótidos, Bantú y Congólidos. Son poblaciones altamente pigmentadas (del moreno claro al negro ébano), piel más gruesa que los leucodermos y más densidad de glándulas sudoríparas, segmentos proximales de las extremidades largos, pilosidad poco densa, cabello corto y crespo, cuerpo siempre esbelto y estatura generalmente alta, forma de la nariz variable (en relación a la temperatura), labios gruesos y evertidos. En general tienen pocos caracteres arcaicos (difieren de los Melanesios); líneas dermopapilares más simples que caucasoides, alta frecuencia de grupo O, Rh+ tipo cDe hata el 90% (2% en blancos).
Existen grupos pigmentados en África, Asia (India) y en Oceanía (Melanesios). Aunque se ha postulado un mismo origen para todos ellos, podría tratarse de convergencia por adaptación climática (Andamaneses). El norte de África está ocupado por grupos de influencia caucasoide y zonas de contacto en el Sáhara (saharianos, tuareg, tebú y etiópidos). Al Sur están las poblaciones negroides. El África subsahariana comprende tanto zonas de sabana, desierto y bosque tropical. Los Pigmeos y los Khoisánidos o Káfridos (Hotentotes) no pueden considerarse poblaciones negras melanodermas. Las poblaciones negras son los Sudánidos, Nilótidos, Bantú y Congólidos. Son poblaciones altamente pigmentadas (del moreno claro al negro ébano), piel más gruesa que los leucodermos y más densidad de glándulas sudoríparas, segmentos proximales de las extremidades largos, pilosidad poco densa, cabello corto y crespo, cuerpo siempre esbelto y estatura generalmente alta, forma de la nariz variable (en relación a la temperatura), labios gruesos y evertidos. En general tienen pocos caracteres arcaicos (difieren de los Melanesios); líneas dermopapilares más simples que caucasoides, alta frecuencia de grupo O, Rh+ tipo cDe hata el 90% (2% en blancos).
- Sudaneses: Sur del Sahara, clima intertropical muy cálido y estacional, al Sur sabana y bosque abierto. Caracteres: cabeza dolicocéfala, frente vertical y estrecha, cara baja, pómulos desarrollados, nariz ancha y deprimida, estatura elevada (1,70 m), pigmentación oscura, pilosidad escasa.
- Nilótidos: Alto Nilo, lagos Rodolfo y Victoria. Caracteres: estatura elevada, longilineos, dolicocéfalos, pigmentación muy oscura, cabello negro, corto y crespo, extremidades largas, cierta variación geográfica de estatura y forma nasal.
- Bantú: lenguas bantú comunes, originarios de los grandes lagos se expandieron hasta Sudáfrica (occidentales, orientales y meridionales).
- Congólidos: paleonégridos, comparten hábitat con pigmeos. Caracteres: estatura media-alta, tronco largo y extremidades cortas, labios menos espesos y piel más clara que congólidos.
- Cáfridos: raza sudafricana, desde Kenia hasta Ciudad del Cabo (cafres, zulús, basutos, suahilis), están muy mezclados, estatura mediana, cuerpo robusto, miembros cortos, espalda y tórax ancho, pigmentación pardo-oscura, dolicocéfalos, son los colonizadores de Madagascar.
- Pigmeos y Khoisánidos: África del Sur, economía típica del Paleolítico Superior, diferenciación temprana de Homo sapiens modernos, prolongado aislamiento, podrían estar relacionados con los pigmeos de Asia (aunque también podría ser una convergencia), actualmente se limitan al desierto del Kalahari (llegaron hace mil años, primero los San, Bosquimanos, y después los Khoi, Hotentotes, empujados por los Bantú).
- Bosquimanos (San): estatura baja (1,55 m), piernas cortas y tronco largo, esteatopigia (depósito de grasa en nalgas sobre todo en mujeres), excelente adaptación al desierto y para el embarazo, piel algo clara parecida a mongoloides, pilosidad escasa, cabello en grano de pimienta, nariz ancha, orejas sin lóbulos, caracteres paidomórficos faciales y craneales.
- Hotentotes (Khoi): estatura algo mayor (1,60 m), piel algo más oscura, suele haber lóbulo en la oreja, nariz menos ancha, esteatopigia aún más acusada.
- Pigmeos: baja estatura (inferior a 1,50 m), pero medio 48 Kg, extremidades inferiores proporcionalmente largas (antebrazo), posteriores cortas (muslo), ventaja adaptativa en ambiente de bosque ecuatorial, pigmentación algo clara, pilosidad corporal abundante, barba poblada.
Variabilidad poblacional caucasoide
Se encuentran en Europa, Norte de África y Asia, y se distinguen básicamente cuatro grupos: 1) poblaciones nórdicas despigmentadas, 2) braquicéfalos de Centroeuropa y Asia Menor, 3) Dolicocéfalos meridionales y 4) formas transmediterráneas.
- Caucasoides Nórdicos: incluye grupos diversos (nórdicos y baltico-oriental).
- Nórdicos: ojos y piel claros, pelo rubio, estatura elevada, nariz estrecha y larga, labios delgados, espalda ancha y pelvis estrecha. Incluye a Teutonórdicos (Noruega, Suecia, Dinamarca, Islandoa. BORJ), Dálonórdico (es más robusto, ‘cromagnon rubio’), Fennonórdico (Rusia septentrional). Son un conjunto heterogéneo adaptado a climas con baja radiación solar.
- Báltico-Oriental: más depigmentados que los nórdicos, cabello rubio claro y liso (lacio), algo más bajos de estatura, poblaciones rusas y Polonia, se extienden hacia Oriente hasta el Pacífico, se mezclan con nórdicos (Finlandia, Paises bálticos, Europa Central y oriente).
- Braquicéfalos Centroeuropeos y de Asia Menor: incluye tres grupos de braquicéfalos morenos (alpinos, dináricos, y anatolios o armenoides).
- Alpinos: braquicefalia marcada, curvo-occipitales, cuerpo rechoncho, estatura media. Probablemente llegaron a Europa en época reciente procedentes del Este.
- Dináricos: braquicéfalos muy marcados, plano-occipitalia, elevada estatura, cara alta y estrecha, platirrinia, pigmentación algo más oscura, iris y pelo pardo oscuro, pilosidad acentuada. Ocupa los Alpes y el Tirol, en otras zonas están mezclados.
- Armenoides: Asia Menor (Siria, Irán, Arabia y hasta la India), braquicéfalo plano-occipital y bóveda alta, cara alta, piel ligeramente morena.
- Dolicocéfalos meridionales: poblaciones mediterráneas, formas gráciles, estatura baja, pigmentación marcada, cabello ondulado o rizado.
- Mediterráneo grácil: caracteres atenuados, dolicocéfalo muy marcado (de la Península Ibérica hasta los Balcanes y el Mediterráneo).
- Mediterráneo robusto: caracteres marcados, mayor estatura y robustez (Portugal, litoral levantino).
- Pirenaico Occidental: constitución robusta, introversión de basion, caracteres serológicos poco comunes (Pirineo Occidental, tipo vasco).
- Formas Transmediterráneas: Ocupan el litoral Norteafricano desde Marruecos hasta Egipto y Canarias. El desierto del Sahara era húmedo al final de la última glaciación (10-7.000 BP). El camello lo introdujeron los romanos (se conquista el desierto). Son poblaciones caucasoides mezcladas, donde predomina el elemento mediterránido (beréberes). Son dolicocéfalos (excepto los Etiópidos). Se pueden agrupar en 5 grupos: Tuareg, Tebú, Saharianos, Etiópidos y Orientálidos.
- Tuareg: proceden de los európidos del Norte de África (mediterránidos) y entran en el desierto, se organizan en castas (vassallos o Imrad, primeros grupos; nobles o ImochagK; y esclavos o Iravelan de clara influencia Sudaneses.
- Tebú: más al Este; parecen ser etiópidos ancestrales pero mezclas entre négridos y európidos (etiópidos), actualmente con una distribución reducida; altos, piel oscura, cabello largo y crespo.
- Saharianos: extremo Atlántico, al Sur del Atlas hasta Senegal; mezclas entre bereberes, con aportes quizá de etiópidos.
- Etiópidos: Desde el Nilo en la 2ª catarata hasta contacto con los Masai (Nilótidos). Mezcla entre európidos (facciones de la cara) y négridos (pirmentados). Región de Nubia, zona de contacto (influencia genética).
- Orientálidos: árabes o semitas (Arabia y Anatolia al Norte, y hasta la India), muy dolicocéfalos, más pigmentados que los mediterráneos (aclimatación?).
- Formas Transasiáticas: Constituyen el complejo racial índido. Desde Iran por el Himalaya hasta Indochina. Influencia európida y sínida (tibetanos del Himalaya). Todo tipo de ambientes, desde desértico al Oeste hasta selvas muy densas al Este. Incluye los grupos de Veddhas, Dravidas, Índidos y mongólidas de la India.
- Veddhas: antiguo y aislado, en sánscrito ‘Veda’ significa ‘Saber’. Aspecto grácil, estatura baja (1,50-1,55 m), tronco ancho, extremidades largas, pómulos acusados, nariz ancha, labios gruesos, algo de prognatismo alveolar, bastante pigmentados. Los Veddhas del Sur están menos mezclados con índidos y son más pigmentados. Particularmente pigmentados son los Veddhas de la Isla de Ceylan que son parecidos a los Australianos. Son cazadores-recolectores, con tecnologías primitivas (relacionados con los primeros humanos modernos salidos de África que llegarían a Australia hace 75.000 años.
- Drávidas: altiplanicies de India central y meridional, representa la población autóctona anterior a la llegada de los blancos. Dolicocéfalos, más altos que Veddhas (1,60 m), piel muy oscura, hasta negra, caracteres faciales robustos, nariz moderadamente ancha, labios carnosos. Dos grupos: Kolariano o Munda (mezclados con indoafgano y mongoloide) y los Tamil (más europoides por su mezcla con los índidos).
- Índidos o indo-afganos: estatura hasta 1,70 m, piel morena más o menos clara, cabello negro y ondulado, rasgos faciales finos, ojos grandes y negros. Se asemejan a los mediterránidos. Origen európido. Toda (parecidos a los Ainú por la pilosidad), Indoario (piel clara), Dardos (estatura mediana) y Singaleses (más pigmentados, Ceylán).
Variabilidad poblacional mongoloide
Son poblaciones ampliamente distribuidas. En la parte Oriental de Asia también hay elementos blancos. Distribución: Siberia, China, Indonesia, Japón. Incluye diversos grupos: Sibéridos (Paleosiberianos), Normongólidos (Túngidos), Centromongólidos (Sínidos), Surmongólidos (Paleomongólidos), Turánidos (turco-tártaros) y Ainú. Caracteres: hay una gran variabilidad (multitud de hábitats). Pigmentación variable (tonos pardos o amarillentos), desde claros a oscuros; cabello lacio, grueso, pilosidad corporal escasa; estatura moderada pero variable; tronco largo respecto a las extremidades; predominan los braquicéfalos; rasgos faciales mongoloides más o menos marcados (malares prominentes, pliegue parpebral ‘ojo mongólido’, raiz nasal hundida, incisivos en pala frecuente, freceuncia elevada de torbellinos digitales.
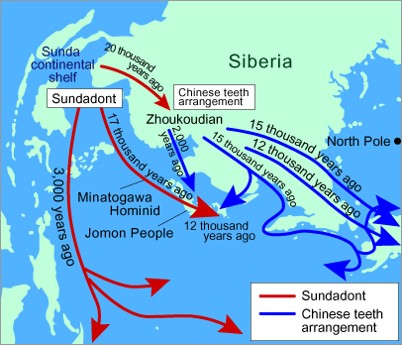
Expansión de las poblaciones Sundadontas y Sinodontas.
- Paleosiberianos: sibérida y uroaltaica; seminómadas de las estepas de Asia Septentrional (al Norte) hasta el estrecho de Behring (esquimales Chukchi, Vogules).
- Túngidos: normongólidos; piel amarilla cobriza, pelo negro y liso, extremidades cortas, cara plana, ojo mongólido marcado (Buriatos, Tunguses estatura 1,55-1,60, Samoyedos tipología atenuada y 1,60 m).
- Sínidos: centromongólidos; Sur de Mongolia hasta y hasta el Tibet, comprende chinos y tibetanos; estatura elevada (hasta 1,70m variable, tibetanos menor); Sherpas (aislados Himalaya, adaptación a la altitud).
- Paleomongólidos: Sudeste Asiático; estatura baja 1,50-1,60 m, pigmentación pardo-amarilla, cuerpo grácil, algo prognatos, ojo mongoloide atenuado. Hacia el Sur del área de distribución los caracteres son más marcados (pigmentación más oscura, más altos, nariz más ancha, cabello ondulado, ojos menos inclinados). Más parecidos a los Veddhas. Los Japoneses son Sínidos migrados desde Korea.
- Turánidos: pastores nómadas del Turkestán ruso; mezclas de contacto mongólidos-caucasoides, cráneo corto, cara alargada.
- Ainú: ocupan el archipiélago japonés, probables descendientes de los Jomones y desplazados pos los Japoneses hacia el Norte. Originalmente procederían de antiguos pobladores de Siberia (descendientes de los primeros Homo sapiens de Asia). Alguna vez se les ha atribuido influencia Európida. Caracteres: gran pilosidad corporal, cabello espeso y rizado, barba (ideal de belleza, factor cultural), cuerpo robusto, baja estatura (1,58 m), ausencia en algunos de pliegue mongólido, piel clara (incluso blanca). Orígen: poblaciones sundadontas actualmente representadas por Australianos, Vedhhas y los Toda hindúes.
- Lapones: zona de contacto troncos európido y mongoloide (formas antiguas y marginales), extremo Norte desde Noruega hasta la antigua U.R.S.S. Estatura baja (1,60 m) piernas cortas, braquicéfalos, pliegue ocular mongoloide en algunos, pómulo prominente. Actualmente se consideran el stock euroasiático más antiguo, habrían ocupado un área más amplia hacia el Sur que en la actualidad.
Variabilidad poblacional ameríndia
Derivan de progresivas migraciones a través del estrecho de Behring. Se relacionan con mongólidos pero algunas migraciones son anteriores al desarrollo de los caracteres de los Sínidos.
 Esquimales: Norte de Alaska y Canadá. Origen hace 7.000 años. Hace 5.000 ya habría ocupado Groenlandia. Son homogéneos en morfología y lengua. Estatura baja (1,60 m), cuerpo robusto, extremidades cortas, pigmentación mongoloide, cráneo y cara mongoloide típica. Cabellos negros y gruesos, pilosidad corporal escasa.
Esquimales: Norte de Alaska y Canadá. Origen hace 7.000 años. Hace 5.000 ya habría ocupado Groenlandia. Son homogéneos en morfología y lengua. Estatura baja (1,60 m), cuerpo robusto, extremidades cortas, pigmentación mongoloide, cráneo y cara mongoloide típica. Cabellos negros y gruesos, pilosidad corporal escasa.- Ameríndios: indígenas americanos en general, descendientes de los primeros pobladores asiáticos hace unos 15.000 años. Estatura media-alta (hasta 1,80 m en patagones y 1,50 en brasílidos). Pigmentación variable pero clara, pilosidad escasa pero variable. Acción de la deriva genética sobre grupos poblacionales pequeños. La cara mongoloide esta atanuada en ameríndios.
- Norpacíficos: (pacífidos y márgidos) Desde Alaska hasta las Rocosas y el Pacífico.
- Sílvidos: noratlántidos (plánidos y apalácidos). Vertiente occidental desde las Rocosas al Atlántico.
- Neoameríndia: surpacífica (puéblidos y Istímidos. Grupos centroamericanos descendientes de las Culturas Mayas y Aztecas.
- Sudatlántica: brasílidos o amazónidos.
- Pámpida: patagones, en franco retroceso o mezcla.
- Paleoameríndia: lágidos y fuéguidos (Selknam, Alakaluf, Yagan).
Variabilidad poblacional de Oceanía
Incluyen las poblaciones de los archipiélagos del Pacífico: Melanesia, Polinesia y Micronesia. También las poblaciones de Australia, Indonesia, Península Malaca, Islas de Andamán, Malasia y las Islas de Andamán.
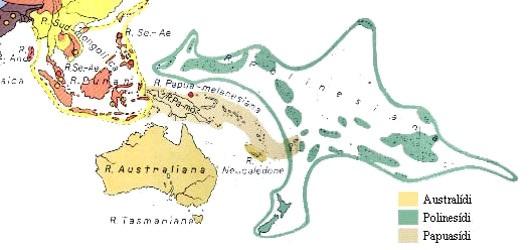
- Archipiélago Malayo: Se distinguen dos conjuntos raciales: negritos y malayos.
- Negritos: en 1521 se produce la llegada de los españoles a Filipinas. Divisaron individuos melanodermos de pequeña estatura en las montañas de Luzón, en la Península de Malaca y en las Islas de Andamán. Etatura 147-150 cm, pigmentación oscura, cabello crespo, nariz ancha pero prominente, pilosidad corporal escasa (difieren de los pigmeos africanos). Se clasifican en Andamaneses (los más aislados y sin mezclar), Seman (viven en junglas de la península de Malaca, bastante mezclados) y Negritos (en lugares montañosos de Filipinas, habrían llegado desde Indochina en el último periodo glacial a través de islas unidas al continente.
- Malayos: incluye la mayor parte de la población Melanesia, poblaciones muy mezcladas. Se distinguen las tribus del interior (dolico o mesocéfalos protomalayos) y las tribus del litoral (braquicéfalos con rasgos mongoloides.
- Protomalayos: baja estatura (155-160 cm, no pigmeos), morenos claros, cabello negro y liso, pómulos pronunciados, labios gruesos, ojos NO mongólidos, nariz frecuentemente aplastada (algo parecidos a los Veddhas, pero cláramente emparentados con los mongoloides), posible mezcla con los negritos que son anteriores.
- Deuteromalayos: población costera de Melanesia y Java, con rasgos más mongoloides, 160-163 cm, cabello liso, nariz ancha y aplastada, ojos oblicuos, morfología grácil. Resultan de la mezcla de indonesios primitivos con elementos mongoles del continente, llegaron a colonizar Madagascar. Los Negritos son cazadores, los protomalayos son agricultores y los deuteromalayor explotan con preferencia recursos marinos.
- Australia: En el momento de su descubrimiento eran unos 300.000, actualmente sólo llegan 80.000. Tienen una morfología primitiva aunque su origen no sería superior a 10-15.000 años. Bastante pigmentados, cabello rizado (nunca crespo), pilosidad abundante, dolicocéfalos, bóveda baja y frente huidiza, arcos superciliares marcados, ojos hundidos, nariz aplastada, cuerpo grácil y esbelto. Se les ha relacionado con Melanesios y Veddhas. No son los más antiguos de Australia.
- Melanesia: Población de piel oscura y cabello crespo parecidos a los “negros de África”. Estatura variable 160-165 cm, piel oscura, cabello crespo pero largo, dolicocéfalos, frente algo huidiza con arcos superciliares algo marcados. Gran variabilidad: Papúes en Nueva Guinea, son de menor estatura; Melanesios,
grupo más reciente, ocupa las otras islas.
Bibliografía
- J. Kingdon (1993) “Self Made Man” John Wiley & Sons Inc.
- Williams Howells (1997) ‘Getting Here’
